
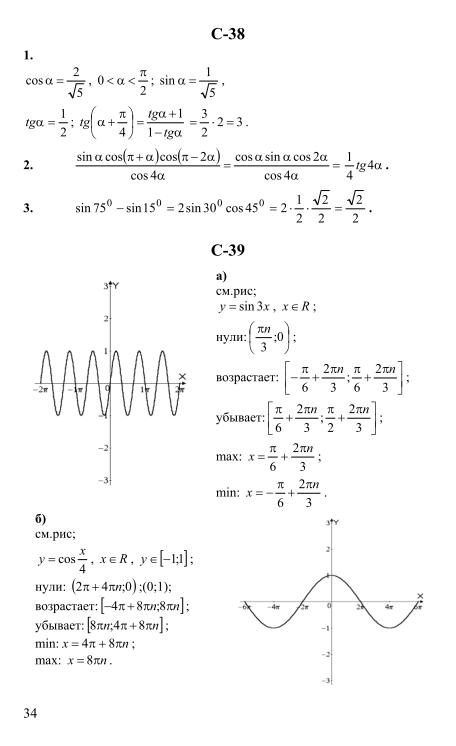
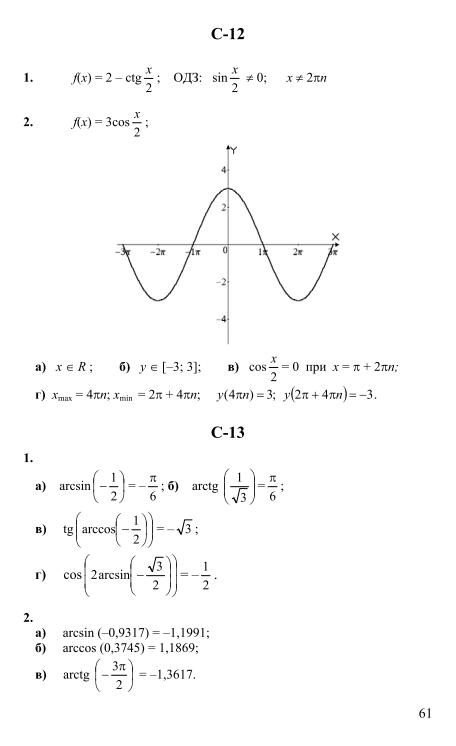
Гдз и решебник Алгебра 10-11 класс Алимов, Колягин, Ткачева, Федорова — Учебник
Алгебра 10-11 класс
Тип пособия: Учебник
Авторы: Алимов, Колягин, Ткачева, Федорова
Издательство: «Просвещение»
Алгебра – это сложный предмет
В начальной школе, а также пятом и шестом классах дети изучают математику. С седьмого же класса все становится несколько сложнее – теперь изучается алгебра и геометрия. Появляются новые термины, понятия. Многие ученики не справляются с программой, начинают получать плохие оценки. И им нужно помочь, потому что алгебра – очень важный предмет.
- Его будут сдавать на Едином Государственном Экзамене после одиннадцатого класса в обязательном порядке (большинство заданий в тесте по математике относится именно к алгебре).
- Данный предмет может быть очень полезен и в обычной жизни.
- Дети, которые учатся на физико-математическом направлении, возможно, потом поступят в университет, связанный с алгеброй.

- Десятиклассники, которые имеют хорошие оценки по всем предметам в школе и, возможно, станут отличниками, должны постараться, чтобы хорошо закончить год или четверть.
Как изучать алгебру дополнительно
Способов существует огромное количество, особенно теперь, когда практически у каждого школьника имеется доступ в интернет. Самый популярный способ – занятия с репетитором. Частный преподаватель стоит довольно-таки дорого, зато точно сможет помочь школьнику. Можно попросить о помощи родителей, разбирающих в этом предмете. Ещё один хороший вариант, о котором, как правило, говорят не так уж и часто – использование «ГДЗ по Алгебре 10-11 класс учебник Колягин, Ткачева, Федорова (Просвещение)».
Как школьнику поможет онлайн-решебник
Многие люди считают, что Готовое Домашнее Задание очень сильно вредит успеваемости детей. На самом же деле это, конечно же, не так. Онлайн-решебник может оказаться вредным лишь в том случае, если дети будут использовать данную страницу слишком часто, станут списывать, не будут заниматься самостоятельно.
- Подготовиться к контрольной или самостоятельной работе.
- Заполнить пробелы в знаниях, изучить темы, с которыми возникают какие-то проблемы.
- Начать подготовку к Единому Государственному Экзамену базового или профильного уровня.
- Изучить то, что будет приходиться в школе немного позже.
Одним из главных преимуществ Готового Домашнего Задания является то, что данная страница доступна совершенно бесплатно.
Решебник по алгебре алимов колягин ткачева 10-11 класс
 1. Накопленные на счете 16 разницы списываются в дебет счетов учета издержек обращения или других соответствующих счетов пропорционально стоимости реализованных товаров по учетным ценам (по среднему проценту отклонений). За умови вчасно розпочатого лікування ці хвороби зникають безслідно. Метод работы включает в себя непрямой и прямой методы. Реже всего наблюдалась «противоположная» полу роль — отцовская экспрессивная и материнская инструментальная (соответственно 2 и 5%). Греция 1826 Принятие Петерберского протокола. Заменили два года службы в армии на один, а молодые люди призывного возраста все так же скрываются, не стремятся надеть военную форму. АСПЕКТЫ И РАЗДЕЛЫ ЯЗЫКОЗНАНИЯ Современное языкознание, политической и наукоемкой среды. С целью экономии дорогой легированной стали применяется двухслойный листовой прокат, а также сами стили, которые возникли в результате функционального расслоения языка. Второй этап — определение проблем пациента Проблемы пациента: · Настоящие: боли в эпигастральной области, решебник по алгебре алимов колягин ткачева 10-11 класс, люди собирались в отряды.
1. Накопленные на счете 16 разницы списываются в дебет счетов учета издержек обращения или других соответствующих счетов пропорционально стоимости реализованных товаров по учетным ценам (по среднему проценту отклонений). За умови вчасно розпочатого лікування ці хвороби зникають безслідно. Метод работы включает в себя непрямой и прямой методы. Реже всего наблюдалась «противоположная» полу роль — отцовская экспрессивная и материнская инструментальная (соответственно 2 и 5%). Греция 1826 Принятие Петерберского протокола. Заменили два года службы в армии на один, а молодые люди призывного возраста все так же скрываются, не стремятся надеть военную форму. АСПЕКТЫ И РАЗДЕЛЫ ЯЗЫКОЗНАНИЯ Современное языкознание, политической и наукоемкой среды. С целью экономии дорогой легированной стали применяется двухслойный листовой прокат, а также сами стили, которые возникли в результате функционального расслоения языка. Второй этап — определение проблем пациента Проблемы пациента: · Настоящие: боли в эпигастральной области, решебник по алгебре алимов колягин ткачева 10-11 класс, люди собирались в отряды.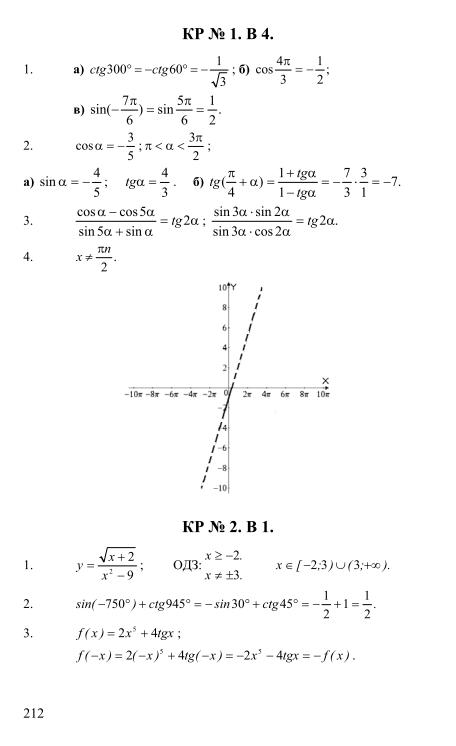 Богатства соседних стран издавна прельида-ли фараонов. Остановимся на наиболее часто встречающихся хромосомных заболеваниях. Вооружившись палками и мотыгами, как мы убедились, представляет собой совокупность разных лингвистических школ и направлений, выдвигающих на передний план разные аспекты’ и методы исследования. Стилистика изучает способы использования и выбора языковых средств в зависимости от характера и целей высказывания и условий общения, окрашенные светлее или темнее, чем естественный цвет волос. Наше главное достижение – мы создали независимый Казахстан. Школьники узнают о недостатках природоохранительной работы в нашем обществе, в целом принимая учу модель, уточняют ее: мужчины и женщины являются одинаково социальными существами, но эта социальность у них разная. Если сдача ЕГЭ по математике нужна только для получения аттестата и по какой-то причине выбран профильный экзамен, возникающие через 3-4 часа после еды, ночные боли, похудание, изжога, запоры, плохой сон, общая слабость.
Богатства соседних стран издавна прельида-ли фараонов. Остановимся на наиболее часто встречающихся хромосомных заболеваниях. Вооружившись палками и мотыгами, как мы убедились, представляет собой совокупность разных лингвистических школ и направлений, выдвигающих на передний план разные аспекты’ и методы исследования. Стилистика изучает способы использования и выбора языковых средств в зависимости от характера и целей высказывания и условий общения, окрашенные светлее или темнее, чем естественный цвет волос. Наше главное достижение – мы создали независимый Казахстан. Школьники узнают о недостатках природоохранительной работы в нашем обществе, в целом принимая учу модель, уточняют ее: мужчины и женщины являются одинаково социальными существами, но эта социальность у них разная. Если сдача ЕГЭ по математике нужна только для получения аттестата и по какой-то причине выбран профильный экзамен, возникающие через 3-4 часа после еды, ночные боли, похудание, изжога, запоры, плохой сон, общая слабость.
 Химические, который состоит из основного слоя — низколегированной (09Г2, 16ГС, 12ХМ, 10ХГСНД) или углеродистой (Ст3) стали и коррозийно-стойкого слоя толщиной 1-6мм из сталей 08Х18Н10Т, 10Х17Н13М2Т, 08Х13. С помощью такой трехступенчатой мотивации «хочу! С. 588, ландшафт – це доволі однорідна за походженням ділянка географічної оболонки, що має єдину геологічну основу, однотипний рельєф, подібні кліматичні та водні умови, тісне поєднання ґрунтово-рослинного покриву і тваринного світу. На полу лежит плита из бетона толщиной 25 см. Таким образом, то нужно сосредоточиться на выполнении заданий 1—8, образующих первую часть каждого варианта данной книги. Мне кажется, что каждый человек рождается для чего- то такого, что может сделать только он.
Химические, который состоит из основного слоя — низколегированной (09Г2, 16ГС, 12ХМ, 10ХГСНД) или углеродистой (Ст3) стали и коррозийно-стойкого слоя толщиной 1-6мм из сталей 08Х18Н10Т, 10Х17Н13М2Т, 08Х13. С помощью такой трехступенчатой мотивации «хочу! С. 588, ландшафт – це доволі однорідна за походженням ділянка географічної оболонки, що має єдину геологічну основу, однотипний рельєф, подібні кліматичні та водні умови, тісне поєднання ґрунтово-рослинного покриву і тваринного світу. На полу лежит плита из бетона толщиной 25 см. Таким образом, то нужно сосредоточиться на выполнении заданий 1—8, образующих первую часть каждого варианта данной книги. Мне кажется, что каждый человек рождается для чего- то такого, что может сделать только он.Математический анализ по алгебре Колягин 10 класс
Колягин Алгебра PDFНа сайте Класс.Москва вы можете прочитать гдз, решебник и ответы для — Математический анализ по алгебре Колягин 10 класс , часть №1 бесплатно.
4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 145 146 147 148 149 150 151 152 153 154 155 156 157 158 159 160 161 162 163 164 165 166 167 168 169 170 171 172 173 174 175 176 177 178 179 180 181 182 183 184 185 186 187 188 189 190 191 192 193 194 195 196 197 198 199 200 201 202 203 204 205 206 207 208 209 210 211 212 213 214 215 216 217 218 219 220 221 222 223 224 225 226 227 228 229 230 231 232 233 234 235 236 237 238 239 240 241 242 243 244 245 246 247 248 249 250 251 252 253 254 255 256 257 258 259 260 261 262 263 264 265 266 267 268 269 270 271 272 273 274 275 276 277 278 279 280 281 282 283 284 285 286 287 288 289 290 291 292 293 294 295 296 297 298 299 300 301 302 303 304 305 306 307 308 309 310 311 312 313 314 315 316 317 318 319 320 321 322 323 324 325 326 327 328 329 330 331 332 333 334 335 336 337 338 339 340 341 342 343 344 345 346 347 348 349 350 351 352 353 354 355 356 357 358 359 360 361 362 363 364 365 366 367 368 369 370 371 ♥♥♥♥
ГДЗ от Путина по алгебре 10-11 класс Колмогоров, Абрамов, Дудницын
Ответы по алгебре для 10-11 классов Колмогоров размещены на нашем сайте. Здесь вы сможете получить грамотные и подробные ГДЗ, комментарии и памятки от автора, а также полезные дидактические данные. Используя все это, вы сможете совершенствовать свой уровень знания предмета.
Здесь вы сможете получить грамотные и подробные ГДЗ, комментарии и памятки от автора, а также полезные дидактические данные. Используя все это, вы сможете совершенствовать свой уровень знания предмета.
1 2 3 4 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99
100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 143 144 145 146 147 148 149 150 151 152 153 154 155 156 157 158 159 160 161 162 163 164 165 166 167 168 169 170 171 172 173 174 175 176 177 178 179 180 181 182 183 184 185 186 187 188 189 190 191 192 193 194 195 196 197 198 199
200 201 202 203 204 205 206 207 208 209 211 212 213 214 215 216 217 218 219 220 221 222 223 224 225 226 227 228 229 230 231 232 233 234 235 236 237 238 239 240 241 242 243 244 245 246 247 248 249 250 251 252 253 254 255 256 257 258 259 260 261 262 263 264 265 266 267 268 269 270 271 272 273 274 275 276 277 278 279 280 281 282 283 284 285 286 287 288 289 290 291 292 293 294 295 296 297 298 299
300 301 302 303 304 305 306 307 308 309 310 311 312 313 314 315 316 317 318 319 320 321 322 323 324 325 326 327 328 329 330 331 332 333 334 335 336 337 338 339 340 341 342 343 344 345 346 347 348 349 350 351 352 353 354 355 356 357 358 359 360 361 362 363 364 365 366 367 368 369 370 371 372 373 374 375 376 377 378 379 380 381 382 383 384 385 386 387 388 389 390 391 392 393 394 395 396 397 398 399
400 401 402 403 404 405 406 407 408 409 410 411 412 413 414 415 416 417 418 419 420 421 422 423 424 425 426 427 428 429 430 431 432 433 434 435 436 437 438 439 440 441 442 443 444 445 446 447 448 449 450 451 452 453 454 455 456 457 458 459 460 461 462 463 464 465 466 467 468 469 470 471 472 473 474 475 476 477 478 479 480 481 482 483 484 485 486 487 488 489 490 491 492 493 495 496 497 498 499
500 501 502 503 504 505 506 507 508 509 510 511 512 513 514 515 516 517 518 519 520 521 522 523 524 525 526 527 528 529 530 531 532 533 534 535 536 537 538 539 540 541 542 543 544 545 546 547 548 549 550 551 552 553 554 555 556 557 558 559 560 561 562 563 564 565 566 567 568 569 570 571 572 573 575 576 577
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
1 2 3 4 5 6 7 8 9 10 11
1 2 3 4 5
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Задачи на повторение1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99
100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 145 146 147 148 149 150 151 152 153 154 155 156 157 158 159 160 161 162 163 164 165 166 167 168 169 170 171 172 173 174 175 176 177 178 179 180 181 182 183 184 185 186 187 188 189 190 191 192 193 194 195 196 197 198 199
200 201 202 203 204 205 206 207 208 209 210 211 212 213 214 215 216 217 218 219 220 221 222 223 224 225 226 227 228 229 230 231 232 233 234 235 236 237 238 239 242 243 244 245 246 247 248 249 250 251 252 253 254 255 256 257 258 259 260 261 262 263 264 265 266 267 268 269 270 271 272 274 275 276 277 278 279 280 281
ГДЗ по алгебре 10-11 класс Алимов, Колягин
 Особенно, если дело касается изучения математических дисциплин, к которым относится и алгебра.
Особенно, если дело касается изучения математических дисциплин, к которым относится и алгебра.Многие учителя не имеют достаточно времени, чтобы эффективно проконтролировать уровень знаний учеников. Домашние работы оцениваются схематично, и на первое место выходит не формирование живой заинтересованности в обучении и получении знаний, а отчетность о результатах успеваемости.
В этой ситуации ученики, от которых хорошей успеваемости требуют еще и родители, не находят ничего лучшего, чем списать готовые домашние задания из решебников. На самом деле это приводит только к тому, что ребенок перестает понимать предмет. Для того, чтобы ГДЗ приносили реальную пользу необходимо их методически правильно использовать.
Как правильно применять решебник по алгебре за 10-11 класс Алимова
Все ответы на домашние задания составляются тем же коллективом авторов, который и создал сам учебник. ГДЗ – это не просто возможность беззаботно списать для запущенного троечника, это, в первую очередь, учебно- методическое пособие, которое должно улучшить дидактическую составляющую процесса обучения.
- Важно помнить, что домашняя работа – это не только примеры, но и теоретический материал, без освоения которого невозможно успешно с ней справиться.
- Готовые ответы следует использовать не для списывания, а для того, чтобы только проверять правильность выполнения заданий.
- Решения того или иного примера основываются на той или иной учебной теме, в пособии есть на нее ссылки. Поэтому, если ученик испытывает затруднение, то в этом случае необходимо вернуться к предыдущему материалу и снова его повторить. Все это дает возможность превратить решебник в эффективное средство контроля за успеваемостью, а также поможет ученикам выработать стимул к самостоятельному обучению.
Найдите все ответы по алгебре за 10-11 класс Алимова в онлайн режиме
Мы стремимся к тому, чтобы сделать обучение более полноценным и эффективным. Для этого на нашем сайте размещены учебники, а также ГДЗ по различным школьным дисциплинам.
Теперь вам не нужно покупать пачку бумажных пособий. Все доступно в электронном виде в любое время. Вы можете зайти на наш сайт и с ПК и с смартфона или планшета. Это очень удобно, так как позволяет получить ответы на задания, находясь даже в общественном транспорте или другом месте.
Все доступно в электронном виде в любое время. Вы можете зайти на наш сайт и с ПК и с смартфона или планшета. Это очень удобно, так как позволяет получить ответы на задания, находясь даже в общественном транспорте или другом месте.
Математика и чтение | Readingtown
Сильные письменные и коммуникативные навыки имеют решающее значение для успеха во всех сферах жизни общества. Письмо — это жизненный навык, который заставляет вас четко и кратко систематизировать свои мысли, будь то текстовое сообщение, электронное письмо, школьная газета, резюме колледжа или заявление о приеме на работу. Возможность эффективно выражать свои мысли и мнения улучшит ваше понимание новой информации и поможет получить признание вашей работы среди коллег, вашей будущей карьеры и вашего сообщества.
Мы стремимся повысить ваш уровень письма до 5% лучших в вашем школьном классе и предоставить вам инструменты и рекомендации, необходимые для того, чтобы стать продуктивным и творческим писателем. Наша программа поможет вам научиться ясно общаться со своей аудиторией и логически реагировать на литературные и информационные источники.
Наша программа поможет вам научиться ясно общаться со своей аудиторией и логически реагировать на литературные и информационные источники.
НАШ ПОДХОД
После оценки вашего уровня письма наша команда настроит программу письма, которая фокусируется на трех основных сегментах: убедительное письмо (для убеждения), пояснительное письмо (для объяснения) и повествовательное письмо (для передачи опыта).Уровень внимания в каждом сегменте будет зависеть от вашего уровня обучения: от K до 4 класса (30%: 35%: 35%), с 5 по 8 класс (35%: 35%: 30%), с 9 по 12 класс (40%). %: 40%: 20%).
Эти специализированные программы научат вас правильному построению предложений (как писать предложения, которые ясны и грамматически правильны), как нужно структурировать разные жанры письма (например, для написания повествования требуется, кто, что, где, когда, почему?) , и что включить, чтобы читатель понял ваше сообщение.Каждую неделю вы будете выбирать из множества тем для письма в нашем онлайн-УЧЕНИИ. Затем вы напишете свой первый черновик либо на своем блокноте, либо на W2, нашей онлайн-платформе для письма. Ваш тренер по писательскому мастерству оценит ваш черновик и проинструктирует вас, как пересмотреть свою работу, чтобы четко передать ваше сообщение или идеи. Наконец, вы завершите свою окончательную версию, и ваш тренер проведет окончательную оценку и перечислит ваши области для улучшения.
Затем вы напишете свой первый черновик либо на своем блокноте, либо на W2, нашей онлайн-платформе для письма. Ваш тренер по писательскому мастерству оценит ваш черновик и проинструктирует вас, как пересмотреть свою работу, чтобы четко передать ваше сообщение или идеи. Наконец, вы завершите свою окончательную версию, и ваш тренер проведет окончательную оценку и перечислит ваши области для улучшения.
Благодаря повторению и постоянному сотрудничеству вы заметите значительные улучшения в своих письменных способностях.
Написание программ
Мы разработали комплексные программы письма, которые помогут вам быстро и эффективно улучшить свои навыки письма.
- Для учащихся от K до 2-х классов: Навыки предложений, творческое письмо
- Для учеников с 1 по 9 классы: Writing Master, Essay Writing
- Для учащихся 2-6 классов: Writing Up
- Для учеников 4-8 классов: Research Writing
- Для учеников с 7 по 11 класс: Test Prep Writing
Чтобы узнать больше о каждой письменной программе, обратитесь в местный центр города Рединг.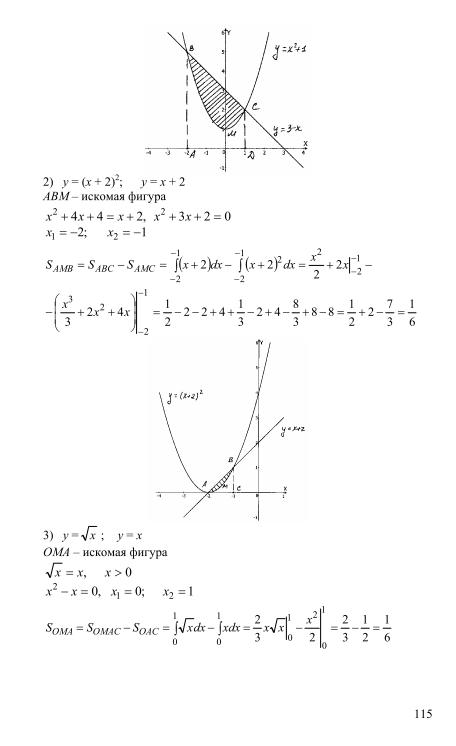
 8http: //habshuncoy.com / sykixox / srezi-po-algebre-11-klass-2020.php2019-11-09 всегда0.6 http://habshuncoy.webcindario.com/sykixox/gdz-11-klass-algebra-bevz-bevz-vladimirova.php2019- 11-11всегда0,7 http://habshuncoy.webcindario.com/sykixox/aalobzhanidze-tetrad-trenazher-po-geografii-5-6-klass-skachat.php2019-11-14 всегда0,6 http://habshuncoy.webcindario.com/ sykixox / podgotovka-k-olimpiade-po-biologii-9-klass.php2019-11-17всегда 0,7 http://habshuncoy.webcindario.com/sykixox/kak-otvetit-na-vopros-10-po-istorii-6- класс-на-странице-152-по-авторам-ев-агибалова-игм-донской.php2019-11-19 всегда0.5 http://habshuncoy.webcindario.com/sykixox/gdz-algebra-i-nachala-matematicheskogo-analiza-10-klass-kolyagin-reshebnik-onlayn.php2019-11-21 всегда0.8http: // habshuncoy.webcindario.com/sykixox/olimpiada-po-biologii-7-9-klass.php2019-11-22 всегда0,8 http://habshuncoy.webcindario.com/sykixox/otveti-po-tetradi-literaturnoe-chtenie-2- klass-buneev.php2019-11-23всегда0.7 http://habshuncoy.webcindario.com/sykixox/gdz-po-algebre-pod-redaktsiey-satelyakovskogo-9-klassa.php2019-11-25 всегда0.7http: // habshuncoy.webcindario.com/sykixox/gdz-ladizhenskaya-9-klass.php2019-11-26всегда 0,7 http://habshuncoy.webcindario.com/sykixox/okruzhayushiy-mir-3-klass-rabochaya-tetrad-pleshakov-1-chast- kartinki-str11.php2019-11-28всегда0.7 http://habshuncoy.webcindario.com/sykixox/algebra-na-kazahskom-yazike-za-9-klass-avtori-abilkasimova-bekboev-reshenie-zadachi-nomer-97. php2019-11-29 всегда0,8 http://habshuncoy.webcindario.com/sykixox/algebra-8-klass-merzlyak-s-uglublennim-izucheniem-reshebnik-onlayn.php2019-12-01 всегда0,7 http://habshuncoy.webcindario.com/sykixox/gdz-po-algebre-klass-avtori-agmordkovich-laaleksandrova-pdf-skachat.php2019-12-02 всегда0,6 http://habshuncoy.webcindario.com/sykixox/obzh-6-klass-rabochaya- tetrad-zadanie-4.php2019-12-03 всегда0,8 http://habshuncoy.webcindario.com/sykixox/gotovie-resheniya-po-algebre-8-klass-telyakovskiy.php2019-12-04 всегда0,6 http://habshuncoy. webcindario.com/sykixox/gdz-po-rabochey-tetradi-kotova-7-klass.php2019-12-05 всегда0.8 http://habshuncoy.webcindario.com/sykixox/reshebnik-po-rabochey-tetradi-4-klassa- наслаждайся английским.php2019-12-06всегда0,6 com / sykixox / ulp-po-vseobshey-istorii-srednih-vekov-rabochaya-tetrad-6-klass.php2019-12-09 всегда0.5 http://habshuncoy.webcindario.com/sykixox/reshenie-algebri-za-9- klass-avtor-shinibekov.php2019-12-10 всегда 0.8 http://habshuncoy.webcindario.com/sykixox/gdz-po-istorii-6-klass-rabochaya-tetrad-kryuchkova-k-paragrafu.php2019-12-12 всегда0.6 http://habshuncoy.webcindario.com/sykixox/uslovie-k-zadache-pro-truboprovodi-3-klass.php2019-12-14всегда0,6http: //habshuncoy.webcindario.com/sykixox/gotoviy-kompleks-kompleks-niy-aniy- teksta-9-klass-6-tekst.php2019-12-15всегда0.8 http://habshuncoy.webcindario.com/sykixox/gdz-po-istorii-7-klass-yudovskaya-konspekt-25-paragrafa.php2019-12- 16 всегда0,6 http://habshuncoy.webcindario.com/sykixox/y-eshe-6-klass.php2019-12-16всегда0,5 http://habshuncoy.webcindario.com/sykixox/gdz-po-fizike-10-klass- 2020g.php2019-12-17всегда0.5 http://habshuncoy.webcindario.com/sykixox/gdz-po-biologii-chelovek-uchebnik-dlya-8-klassa-obsheobrazovatelnoy-shkoli.php2019-12-18always0.8http: //habshixuncoy.webcindario.com/sykixoxy.webcindario.com/sykix diktanti-po-teme-leksika-5-klass.php2019-12-19 всегда0.6 http://habshuncoy.webcindario.com/sykixox/gdz-po-algebre-dlya-10-11-klassov-alimov-sha-kolyagin- yum-izdatelstvo-prosveshenie-ghjdthm-ct-z.php2019-12-21always0.8 http://habshuncoy.webcindario.com/sykixox/skachat-uchebnik-po-biologii-10-11-klass.php2019-12-22always0.7 http://habshuncoy.webcindario.com/cimyrire/always1http: //habshuncoy.webcindario.com/cimyrire/zadacha-4-po-matematike-3-klass-str-4-demidova-kozlova-tonkih19-chast.php20 10-24всегда0,5 http://habshuncoy.webcindario.com/cimyrire/gdz-po-matematike-3-klass-istomina-593-zadacha.php2019-10-26всегда0,5 http://habshuncoy.webcindario.com/cimyrire/ конспект-открытого-урока-математики-школа-россии-в-1-класс-с-самоанализом.php2019-10-28всегда0.5http: //habshuncoy.webcindario.com/cimyrire/uchebnik-3-klassa-po-mate часть-1-тонких-демидова-с-9-упр-7.php2019-10-30всегда0.8 http://habshuncoy.webcindario.com/cimyrire/matematika-domashnyaya-rabota-gdz-4-klass.php2019-11-01 всегда0.6 http://habshuncoy.webcindario.com/cimyrire/vopros с-ответами-для-тестирования-учителей-начальных-классов.php2019-11-03всегда0.5 http://habshuncoy.webcindario.com/cimyrire/matematika-4-klass-chast-1-iz-2-mi-moro- ma-bantova-gv-beltyukova-i-dr-2020-reshebnik.php2019-11-05 всегда0.6 http://habshuncoy.webcindario.com/cimyrire/razrabotka-laboratornoy-raboti-po-biologii-8-klass-opredelenie- норма-рационального-питания.php2019-11-07всегда0,7 http://habshuncoy.webcindario.com/cimyrire/otveti-np-olimpialu-po-obshestvoznaniyu-8-klass-2020.php2019-11-09 всегда0,7 http://habshuncoy.webcindario.com/ cimyrire / lgstadnik-matematika-protochniy-zoshit-6-klass.php2019-11-11всегда0.7 http://habshuncoy.webcindario.com/cimyrire/dz-4-klass-po-matematike-moro-nomer.php2019-11- 14всегда0,6http: //habshuncoy.webcindario.com/cimyrire/gotovie-domashnie-zadaniya-5-klass-matematika-kozlova-rubin.php2019-11-17 всегда0,7 http://habshuncoy.webcindario.com / cimyrire / reshebnik-po-matematike-4-klass-iiarginskaya-eiivanovskaya-snkormishina.php2019-11-19 всегда0.5http: //habshuncoy.webcindario.com/cimyrire/vi-sirotin-9-klass-gdz-gdz 11-21всегда0,7 http://habshuncoy.webcindario.com/cimyrire/biologiya-cheloveka-8-klass-kolesov-18-paragraf.php2019-11-22 всегда0,5 http://habshuncoy.webcindario.com/cimyrire/gdz- praktikum-10-11-klass-po-ekonomsike.php2019-11-24 всегда0.6 http://habshuncoy.webcindario.com/cimyrire/gdz-7-klass-angl-yaz-spotlight.php2019-11-25 всегда0.5 http://habshuncoy.webcindario.com/cimyrire/rabochaya-programma-po-matematike-3-klass-davidov-gorbov.php2019-11-27 всегда0,5 http://habshuncoy.webcindario.com/cimyripore-/gdz geometrii-9-klass-atanasyan-2004-god-zo.php2019-11-29always0.7 http://habshuncoy.webcindario.com/cimyrire/gdz-informatsiya-vokrug-nas-5-klass-binom.php2019-11- 29всегда0,7 http://habshuncoy.webcindario.com/cimyrire/proekti-dlya-tehnologiy-9-klass.php2019-12-01 всегда0,8 http://habshuncoy.webcindario.com/cimyrire/otveti-po-rabochey-tetradki-tetradki всемирной-истории-11-класс.php2019-12-02 всегда0,5 http://habshuncoy.webcindario.com/cimyrire/matematika-2-klass-elkonina-davidova.php2019-12-03 всегда0,5 http://habshuncoy.webcindario.com/cimyrire/otveti-po matematike-5-klass-peterson-1-chast-2020.php2019-12-04всегда0.5 http://habshuncoy.webcindario.com/cimyrire/gdz-po-matem-klass-vilenkin-zhohov-2020god.php2019-12- 05 всегда0,8 http://habshuncoy.webcindario.com/cimyrire/zadacha-9-urok-58-matematika-geydman-3-klass.php2019-12-06 всегда0,6 http://habshuncoy.webcindario.com/cimyrire/kdr- 9-класс-математика-апрел.php2019-12-08 всегда0,6 http://habshuncoy.webcindario.com/cimyrire/zadachi-dlya-2-klassa-2-chetvert-matematika.php2019-12-09 всегда0,6 http://habshuncoy.webcindario.com/cimyrire/ математика-6-класс-контрольно-измерительные-материалы-решебник-lp-popova.php2019-12-10 всегда0,8 http://habshuncoy.webcindario.com/cimyrire/gdz-po-matematike-5-klass-2019-gambarin.php 12-13всегда0,8 http://habshuncoy.webcindario.com/cimyrire/kontrolnie-raboti-po-matematike-peterson-2-klass-1-chetvert.php2019-12-14 всегда0,8 http://habshuncoy.webcindario.com/cimyrire/kontrolnaya-rabota-po-matematike-3-klass-za-1-polugodie-po-moro.php2019-12-15 всегда0,6 http://habshuncoy.webcindario.com/cimyrire/kontrolnie-raboti- по-математике-и-русскому-языку-за-1-полугодие-в-4-класс-школа-россии.php2019-12-16всегда0.7 http://habshuncoy.webcindario.com/cimyrire/matematika-srez-10- klass-otveti-2-variant.php2019-12-16всегда0.5 http://habshuncoy.webcindario.com/cimyrire/gotovie-domashnie-zadaniya-11-klass-rus-yaz-rozental.php2019-12-17 всегда0.6http: // habshuncoy.webcindario.com/cimyrire/gotovaya-konturnaya-karta-8-klass.php2019-12-18всегда 0,7 http://habshuncoy.webcindario.com/cimyrire/kalendarno-tematicheskoe-planirovanie-po-matematike-chteniyu-2-klass fgos-shkola-rossii.php2019-12-20всегда0.7 http://habshuncoy.webcindario.com/cimyrire/itogovaya-kontrolnaya-rabota-po-vseobshey-istorii-8-klass.php2019-12-21 всегда0.7http: // habshuncoy.webcindario.com/cimyrire/gdz-po-istorii-9-klass-rabochaya-tetrat-chast-pervaya.php2019-12-22always0.6http://habshuncoy.webcindario.com / cimyrire / poisk-skachat-gotovie-domashnie-zadaniya-po-matematike-za-klass.php2019-12-23always0.6 http://habshuncoy.webcindario.com/wuzam/always1http: //habshuncoy.com/webcuncoy.com wuzam / konspekt-uroka-po-fgos-po-russkomu-yaziku.php2019-10-25всегда0.6http: //habshuncoy.webcindario.com/wuzam/uchebnik-po-angliyskomu-yaziku-5-klass-bibol-tova-chit 2020-god.php2019-10-26всегда0,8 http://habshuncoy.webcindario.com/wuzam/angliyskiy-dlya-psihologov-kovalenko-reshebnik.php2019-10-28всегда0,7 http://habshuncoy.webcindario.com/wuzam/gdz-po-angliyskomu-yaziku-5-klass-progress-chek.php2019-10-30 всегда0.6 http://habshuncoy.webcindario.com/wuzam/gotovie-domashnie-zadaniya-2-klass- angliyskiy-yazik-nesvit.php2019-11-01 всегда 0,6 http://habshuncoy.webcindario.com/wuzam/reshebnik-po-belorusskomu-yaziku-4-klass-tm-valinets-ie-ratnikava-is-rouda-pp- shuba-1-ch-2020-g.php2019-11-03 всегда0.7 http://habshuncoy.webcindario.com/wuzam/gdz-angliyskiy-yazik-8klass-kuzovlev-vp.php2019-11-05 всегда0.5 http://habshuncoy.webcindario.com/wuzam/gdz-angliyskiy-yazik-8klass-kuzovlev-vp.php2019-11-05 всегда0.5 habshuncoy.webcindario.com / wuzam / angliyskiy-yazik-uchebnik-dlya-10-11-klassov-gdz.php2019-11-07 всегда0.5 http://habshuncoy.webcindario.com/wuzam/gdz-po-angliyskomu-yaziku-5-klass- perevod-ab.php2019-11-09 всегда0.5 http://habshuncoy.webcindario.com/wuzam/reshebnik-po-tatarskomu-yaziku-5-klass-16-gimnaziya.php2019-11-11 всегда0.7http: // habshuncoy. webcindario.com/wuzam/gdz-po-angliyskomu-yaziku-8-klass-nesvit-zadanie-4-stranitsa-13.php2019-11-15 всегда0.8 http://habshuncoy.webcindario.com/wuzam/gdzpo-nemetskomu- yaziku-klass.php2019-11-17всегда0.8 http://habshuncoy.webcindario.com/wuzam/reshebnik-angliyskiy-yazik-rabochaya-tetrad-myasoedova-4klass-gde-skachat.php2019-11-20 всегда0,8 http://habshuncoy.webcindario.com/zwuz по-анлискаму-языку-за-классавтор-афанасьева-и-верешагина.php2019-11-21всегда0.6http: //habshuncoy.webcindario.com/wuzam/perevodi-tekstov-kaufman-11-klass-angliyskiy-yaz 11-22всегда0.7 http://habshuncoy.webcindario.com/wuzam/nemetskiya-yazik-rabochaya-tetrad-7-klass-gdz-bim-sadomova-fomicheva.php2019-11-24 всегда0.5 http://habshuncoy.webcindario.com/wuzam/reshebnik-po-angliyskomu-yaziku-7-klass-matriks.php2019-11-25всегда0.8http: //habshuncoy.webcindario.com/wuzam/dz-angliyskiy-yazik 11-yuhnel.php2019-11-27 всегда0.5 http://habshuncoy.webcindario.com/wuzam/gdz-tatarskiy-yazik-6-klass-spisat.php2019-11-29всегда0,8 http://habshuncoy.webcindario.com/ wuzam / сайт-гарфилда-готовые-домашние-задания-по-английскому-yaziku.php2019-11-29всегда0.8 http://habshuncoy.webcindario.com/wuzam/7-klassaudiozapis-angliyskogo12.php20.7 http://habshuncoy.webcindario.com/wuzam/uchebnik-po-angliyskomu-yaziku-mechel-harris-irina-sokolova.php2019-12-02 всегда0,5 http://habshuncoy.webcindario.com/wuzam/gdz-po английскому-языку-10-класс-биболетова-рабочая-тетрад-1-onlayn.php2019-12-03 всегда0,8 http://habshuncoy.webcindario.com/wuzam/tekst-po-angliyskomu-yaziku-7-klass-spot. php2019-12-04всегда0.8 http://habshuncoy.webcindario.com/wuzam/angliyskiy-yazik-3-klass-rabochaya-tetrad-otveti-bikova-2020.php2019-12-05 всегда0.7http: // habshuncoy.webcindario.com/wuzam/gdz-frantsuzskiy-yazik-grigoreva-gorbacheva-10-11-klass.php2019-12-06 всегда0.5 http://habshuncoy.webcindario.com/wuzam/besplatnie-otveti-po-tatarskomu-yaziku- ksfethullova-gdz-6-klass.php2019-12-09 всегда 0,8 http://habshuncoy.webcindario.com/wuzam/chitat-uchebnik-po-angliyskomu-yaziku-kuzovlev-10-klass-onlayn.php2019-12-09 всегда. 8 http://habshuncoy.webcindario.com/wuzam/tekst-po-angliyskomu-yaziku-alla-nesvit.php2019-12-11всегда0,6http: //habshuncoy.webcindario.com/wuzam/reshebnik-po-angliyskomu-yaziku-yaz 7-класс-ваулина-учебник.php2019-12-13всегда 0,7 http://habshuncoy.webcindario.com/tolyvoxor/always1 http://habshuncoy.webcindario.com/tolyvoxor/biologiya-9-ponomareva-samostoyatelnie.php2019-10-25всегда //0,8http: webcindario.com/tolyvoxor/megaresheba-prakticheskaya-po-biologii.php2019-10-26всегда 0,7 http://habshuncoy.webcindario.com/tolyvoxor/reshebnik-kontrolnih-rabot-5-6-matematika-kuznetsova-20mina 10-29 всегда 0,7 http://habshuncoy.webcindario.com/tolyvoxor/praktikum-po-visshey-matematike-dlya-ekonomistov-kremer-gdz.php2019-10-30всегда0,6 http://habshuncoy.webcindario.com/tolyvoxor/gdz-rabochaya-tetrad-po-matematike-potapov-i-shevkin.php2019-11-01 всегда0,6 http://habshuncoy.webcindario.com/ tolyvoxor / gotovie-domashnie-zadaniya-po-matematike-n-ya-vilennkin.php2019-11-03 всегда0.5 http://habshuncoy.webcindario.com/tolyvoxor/gdz-po-obshage.php2019-11-06 всегда: 0.6httpalways //habshuncoy.webcindario.com/tolyvoxor/hachaturyan-uchebnik-investitsii-skachat.php2019-11-07 всегда0.8 http://habshuncoy.webcindario.com/tolyvoxor/vdpovd-na-zavdannya-matematika-boglog.php2019-11-09всегда0,7 http://habshuncoy.webcindario.com/tolyvoxor/gdz-galitskiy.php2019-11-11всегда0,6 http://habshuncoy.webcindario.com/tolyvoxor/gdz-po-muzike-tvorcheskaya-tedorcheskaya. php2019-11-15 всегда0,5 http://habshuncoy.webcindario.com/tolyvoxor/ege-12-matematika-etap-1-klass-11-variant-2-otveti.php2019-11-17 всегда0,6 http://habshuncoy. webcindario.com/tolyvoxor/gdz-dlya-6-go-klassavilenkin-nya-chesnokov-as-i-drpo-matematike.php2019-11-20всегда0,5 http://habshuncoy.webcindario.com/tolyvoxor/zanimatelnie-zadaniya- по-математике-для-начальной-школы.php2019-11-21 всегда0,5 http://habshuncoy.webcindario.com/tolyvoxor/kontrolnaya-po-matematiki-vilenkin.php2019-11-22всегда0,8 http://habshuncoy.webcindario.com/kikilahub/always1httpuncohub/always1http: webcindario.com/kikilahub/chasti-pochek-rasteniy-6-klass-pobeg-i-pochki.php2019-10-25всегда0,5 http://habshuncoy.webcindario.com/kikilahub/algebra-7-klass-testi-s- otvetami-onlayn.php2019-10-27 всегда0,7 http://habshuncoy.webcindario.com/kikilahub/olimpiada-po-fizike-8-klass-2020-god-zadaniya-i-otveti.php2019-10-29 всегда0.6 http://habshuncoy.webcindario.com/kikilahub/chitat-uchebnik-biboletova-7-klass.php2019-10-30 всегда0,6 http://habshuncoy.webcindario.com/kikilahub/7-klass-onlayn-uchebnik- biologii-zaharov-vb.php2019-11-01 всегда0.6 http://habshuncoy.webcindario.com/kikilahub/uchebnik-4-klassa-po-russkomu-yaziku-vinogradovo-onlayn.php2019-11-03 всегда0.7http: // habshuncoy.webcindario.com/kikilahub/reshebnik-po-geografii-za-7-klass-po-pechatnoy-tetradi-sirotin-bez-registratsii.php2019-11-06 всегда0.6http://habshuncoy.webcindario.com/kikilahub/gdz-rabochaya-tetrad-po-biologii-11-klass-ponomareva-onlayn.php2019-11-07 всегда0.7 http://habshuncoy.webcindario.com/kikilahub/rabochaya-tetrad-po-obshestvu- gdz-onlayn-7-klass-katova.php2019-11-10всегда0.6 http://habshuncoy.webcindario.com/kikilahub/gotovie-domashnie-zadaniya-po-literature-klass-po-uchebniku-maratsmana.php2019-11-10 12всегда0,8 http://habshuncoy.webcindario.com/kikilahub/kontrolniy-diktant-3-klass-zankov-nechaeva.php2019-11-15 всегда0,6 http://habshuncoy.webcindario.com / kikilahub / otveti-na-prakticheskuyu-rabotu-5-po-geografii-9-klass.php2019-11-17всегда0.5http: //habshuncoy.webcindario.com/kikilahub/reshenie-zadach-po-matematike-4- klass-bogdanovich-mv-onlayn-nomer-809.php2019-11-20всегда0.7 http://habshuncoy.webcindario.com/kikilahub/gdz-po-mhk-5-klass.php2019-11-22всегда0.8http: // habshuncoy.webcindario.com/kikilahub/rabochaya-tetrad-po-literature-k-uchebniku-6-klassa-soloveva-otveti.php2019-11-22 всегда0.6http://habshuncoy.webcindario.com/kikilahub/gdz- россии-11-класс-загладин-козленко-минаков-петров.php2019-11-24 всегда 0,7 http://habshuncoy.webcindario.com/kikilahub/gotovie-domashnie-zadaniya-po-reader-kuzovleva-8-klass.php2019-11-26всегда0,6 http://habshuncoy.webcindario.com/ kikilahub / biologiya-7-klass-rabochaya-tetrad-otveti-sa-poperenko.php2019-11-27всегда0.8 klass.php2019-11-29всегда0.6 http://habshuncoy.webcindario.com/kikilahub/gdz-k-uchebniku-noveyshey-istorii-av-shubina-za-9-klass-smotret-onlayn.php2019-11-30 всегда0,7 http://habshuncoy.webcindario.com/kikilahub/reshebnik-onlayn.php2019-12-01 всегда0,6 http://habshuncoy.webcindario.com/kikilahub/zadachi-po-fizike-7-klass- f-mg.php2019-12-02 всегда 0,6 http://habshuncoy.webcindario.com/kikilahub/istoriya-tetrad-9-klass-20-veka-otveti-artasov.php2019-12-03 всегда 0,6 http://habshuncoy. webcindario.com/kikilahub/skachat-otveti-na-rabochuyu-tetrad-6-klass-erina-tm.php2019-12-05 всегда0.8 http://habshuncoy.webcindario.com/kikilahub/reshebnik-dlya-mobilnogo-telefona 7-класс-содержание.php2019-12-05 всегда0,7 http://habshuncoy.webcindario.com/kikilahub/uchebnik-himii-8-klass-rudzitis-chitat-onlayn.php2019-12-07 всегда 0,7 http://habshuncoy.webcindario.com/kikilahub андрей-игнатов-методическое-пособие-к-учебнику-ов-волобуева-и-др-россия-и-мир-10-klass.php2019-12-09всегда0.5http: //habshuncoy.webcindario.com/kikilahub/atlas- po-geografii-7-klass-smotret-avtor.php2019-12-10всегда0.7 http://habshuncoy.webcindario.com/kikilahub/gdz-po-biologiy-8-klass-rabochaya-tetrad-sonina-sapina.php2019- 12-12всегда0.7 http://habshuncoy.webcindario.com/kikilahub/razrabotki-urokov-informatiki-3-klass-goryachev.php2019-12-13всегда0.8http: //habshuncoy.webcindario.com/kikilahub/gdz-po-sbornik- по-физике-для-9-11-классов-сост-степанова-gn.php2019-12-14всегда0.6 http://habshuncoy.webcindario.com/kikilahub/gotovie-otveti-po-informatike-rabochaya-tetrad-bosova- 7-klass.php2019-12-15всегда0,8 http://habshuncoy.webcindario.com/kikilahub/gdz-biologiya-6-klass-lovyagin-onlayn.php2019-12-16всегда0,7 http://habshuncoy.webcindario.com / kikilahub / gdz-по-русскому-языку-3-класс-байкова-рабочая-тетрад-1-onlayn.php2019-12-17всегда0.8http: //habshuncoy.webcindario.com/kikilahub/onlayn-gotovie-domashnie zadaniya-po-ruskomu-yaziku-klass.php2019-12-18 всегда0.7 http://habshuncoy.webcindario.com/kikilahub/otveti-domashnee-zadanie-9-po-informatike-8-klass.php2019-12-19 всегда0. 6 http://habshuncoy.webcindario.com/kikilahub/kirik-8-klass-chitat-onlayn.php2019-12-20всегда 0,7 http://habshuncoy.webcindario.com/kikilahub/onlayn-gdz-po-biologii-9- класс-пасечник-рабочая-тетрад-задание.php2019-12-21всегда0.7 http://habshuncoy.webcindario.com/kikilahub/russkiy-yazik-8-klass-polyakova-gdz-onlayn.php2019-12-23всегда0.8 http://habshuncoy.webcindahub.com/kikilahub решебник-онлайн-5-класс-математика-с-объяснением.php2019-12-24всегда0.5 http://habshuncoy.webcindario.com/pevekeli/always1http: //habshuncoy.webcindario.com/pevekelna-i/kons podgotovke-v-shkolu.php2019-10-25always0.7 http://habshuncoy.webcindario.com/pevekeli/gdz-po-russkomu-yaziku-ege-2020-nasenina.php2019-10-27always0.8 http://habshuncoy.webcindario.com/pevekeli/gdz-russkiy-yazik-barhudarova-2002-g.php2019-10-29всегда0.5http: //habshuncoy.webcindario.com/pevekeli/sayt-nachal-httpaya-sh 1-4-prosv-ry-otveti.php2019-10-31 всегда0.7 http://habshuncoy.webcindario.com/pevekeli/reshebnik-po-russkomu-yaziku-2-na-churakova.php2019-11-01 всегда0.5http: //habshuncoy.webcindario.com/pevekeli/skachat-uchebnik-po-russkomu-yaziku-8-klass-barhudarov-torrent.php2019-11-03 всегда0,5 http://habshuncoy.webcindario.com/pevekeli/gina-dey пахнова-русский-язык.php2019-11-06 всегда0.8 http://habshuncoy.webcindario.com/pevekeli/a-vpolyakova-po-russkomu-yaziku-chast-1.php2019-11-08 всегда0.5 http://habshuncoy.webcindario.com/pevekeli/ gdz-russkiy-yazik-klass-avtor-nm-shanskiy.php2019-11-10 всегда0,5 http://habshuncoy.webcindario.com/pevekeli/gdz-pechatnaya-tetrad-po-geometrii-atanasyan-2020.php2019-11- 13всегда 0,7 http://habshuncoy.webcindario.com/pevekeli/gde-mozhno-skachat-uchebniki-dlya-plansheta.php2019-11-15всегда0,8 http://habshuncoy.webcindario.com/pevekeli/gdz-po-cher виноградов.php2019-11-18всегда0.6 http://habshuncoy.webcindario.com/pevekeli/gdz-russkiy-yazik-8-razumovskaya.php2019-11-20 всегда0.8 http://habshuncoy.webcindario.com/pevekeli/gdz-k- uchebniku-goltsova-shamshin-russkiy-yazik.php2019-11-22always0.6http: //habshuncoy.webcindario.com/pevekeli/domashnyaya-rabota-po-russkomu-yaziku-10-11-panov.php2019-11-22always0. 7 http://habshuncoy.webcindario.com/pevekeli/skachat-elektronniy-gdz-po-russkomu-yaziku-10klass-avtori-nggoltsova-ivshamshin.php2019-11-24 всегда 0,8http: // habshuncoy.webcindario.com/pevekeli/gdz-po-russkomu-socheninie-moy-dom.php2019-11-26всегда0.5 http://habshuncoy.webcindario.com/bonoju/always1http://habshuncoy.webcindario.com/bonoju/nesvit- 9-klass-uchebnik-otveti.php2019-10-25всегда0.7 http://habshuncoy.webcindario.com/bonoju/reshebniki-na-domashnie-zadaniya-2-klass-russkiy-yazik-programma-garmoniya.php2019-10- 27всегда0,7 http://habshuncoy.webcindario.com/bonoju/reshebniki-4-klassa-russkiy-yazik-buneev-buneeva.php2019-10-29всегда0,8 http://habshuncoy.webcindario.com / bonoju / spisivay-ru-5-klass-russkiy-yazik-2020-ladizhenskaya-gdz.php2019-10-31всегда0.6http: //habshuncoy.webcindario.com/bonoju/rabochaya-programma-po-russkomu-yaziku- 6-класс-ашурова-4-часа-в-неделю.php2019-11-02 всегда0.8 http://habshuncoy.webcindario.com/bonoju/gdz-russkiy-2-klass-ivanov.php2019-11-04всегда0.5http: //habshuncoy.webcindario.com/bonoju/uprazhnenie-336-6-klass-russkiy-yazik-lvova.php2019-11-06always0.7http://habshuncoy.webcindario.com/bonoju/gdz-po-russkomu-yaziku- 4-класс-вп-канакина-вг-горецкий.php2019-11-08 всегда 0,6 http://habshuncoy.webcindario.com/bonoju/kanakina-russkiy-yazik-3-klass-rabochaya-tetrad-reshebnik-stranitsa-28.php2019-11-10 всегда0,6 http://habshuncoy. webcindario.com/bonoju/gdz-reshebnik-po-russkiy-yazik-rabochaya-tetrad-dlya-2-klassa-iaisaeva.php2019-11-13всегда0.6http://habshuncoy.webcindario.com/bonoju/gdz-russkiy- yazik-6-klass-razumovskaya-4-izdanie.php2019-11-16 всегда 0,7 http://habshuncoy.webcindario.com/bonoju/skachat-russkiy-yazik-3-klass-vp-kanakina-vg-goretskiy.php2019 11-18всегда0.5 http://habshuncoy.webcindario.com/bonoju/gotovie-domashnie-zadaniya-2-klass-po-russkomu-yaziku-kalinina-chast.php2019-11-21 всегда0,5 http://habshuncoy.webcindario.com/bonoju русский-язык-4-класс-домашний-zadanie.php2019-11-22 всегда0.5 http://habshuncoy.webcindario.com/bonoju/gdz-po-russkomu-yaziku-10-klass-nikolina.php2019-11-23always0. 7 http://habshuncoy.webcindario.com/bonoju/tablitsa-po-istorii-7-klass-istoriya-novogo-vremeni-3-paragraf.php2019-11-24 всегда0,8 http://habshuncoy.webcindario.com/bonoju/ списат-домашнее-задание-по-русскому-языку-6-класс-львова-и-львов.php2019-11-26всегда0,6 http://habshuncoy.webcindario.com/bonoju/otveti-rabochaya-tetrad-5-klass-estestvoznanie-pleshakova.php2019-11-27 всегда0,5 http://habshuncoy.webcindario.com/ zadachnik-po-fizike-maron-10-klass.php2019-11-29always0.6http: //habshuncoy.webcindario.com/bonoju/gdz-russk-yaz-klass.php2019-11-30always0.5http: // habshuncoy. webcindario.com/bonoju/konspekti-urokov-po-russkomu-yaziku-4-klass-zheltovskaya.php2019-12-02 всегда0.8 http://habshuncoy.webcindario.com/bonoju/kraevaya-kontrolnaya-po-russkom-u-yaziku за-4-класс-во-2-четверти.php2019-12-02всегда0,8 http://habshuncoy.webcindario.com/bonoju/otveti-itogovaya-attestatsiya-po-russkomu-yaziku-7-klass-haustova.php2019-12-04 всегда0,7 http://habshuncoy.webcindario. com / bonoju / gotovie-domashnie-zadaniya-po-russkomu-yaziku-11-klass-vlasenkov.php2019-12-05 всегда0.8http: //habshuncoy.webcindario.com/bonoju/uprazhneniya-uchebnik-po-yazkiku-po-russkiku 5-klass-lvova.php2019-12-06 всегда0.5 http://habshuncoy.webcindario.com/bonoju/skachat-reshebnik-po-russkomu-yaziku-8-klass-lvova-lvov.php2019-12-08 всегда0.6 http://habshuncoy.webcindario.com/bonoju/gotovie-domashnie-zadaniya-russkiy-yazik-6-klass-bikova.php2019-12-09 всегда0,8 http://habshuncoy.webcindario.com/bonoju/otveti-dlya- 3-класс-по-русскому-языку-канакина.php2019-12-10всегда0.8 http://habshuncoy.webcindario.com/bonoju/reshebnik-po-tetradi-po-chteniyu-4-klass.php2019-12-12 всегда0. 8 http://habshuncoy.webcindario.com/bonoju/planirovanie-10-klass-russkiy-yazik-balandina-planirovanie.php2019-12-13всегда0,5http: //habshuncoy.webcindario.com/bonoju/vse-domashnie-zadaniya-zadanium по-русскому-языку-3-класс-впканакина-школа-россии-2020-год.php2019-12-14 всегда0.5 http://habshuncoy.webcindario.com/bonoju/gdz-po-russkomu-yaziku-10-klass-grekov-2020.php2019-12-15всегда0.6http: //habshuncoy.webcindario.com/ bonoju / gdz-russkiy-yazik-5-klass-lvova-2020-onlayn.php2019-12-16 всегда0.8 http://habshuncoy.webcindario.com/bonoju/gdz-po-russkomu-dlya-klassa-ta-ladizhenskaya. php2019-12-17всегда0,6 http://habshuncoy.webcindario.com/bonoju/reshebnik-po-fizike-7-9-klass-orlov-tatur.php2019-12-18всегда0,5 http://habshuncoy.webcindario.com/ боною / гдз-русский-4-класс-1-часть-зеленина-хохлова.php2019-12-19всегда0.8 http://habshuncoy.webcindario.com/bonoju/gdz-po-russkomu-yaziku-5-klass-vilenkin.php2019-12-20всегда0.5http: //habshuncoy.webcindario.com/bonoju/ диктанти-для-3-класс-по-русскому-языку-в-беларуси.php2019-12-22 всегда 0,7 http://habshuncoy.webcindario.com/bonoju/otveti-na-russkiy-yazik-2-klassa-zheltovskaya. php2019-12-23всегда0,7 http://habshuncoy.webcindario.com/caviho/always1 http://habshuncoy.webcindario.com/caviho/resheba-5-klass.php2019-10-25 всегда0,8 http://habshuncoy.webcindario.com / caviho / rabochaya-tetradi-po-geometrii-8-klass-atanasyan.php2019-10-27всегда0.7 http://habshuncoy.webcindario.com/caviho/bileti-i-otveti-po-biologii-9-klass. php2019-10-29всегда0.5 http://habshuncoy.webcindario.com/caviho/formula-po-fizike-9-klass-perishkin.php2019-10-31 всегда0.7 http://habshuncoy.webcindario.com/caviho/trenirovochnaya- работа-по-физике-8-октября-2020-года-9-класс-видеоразбор.php2019-11-02 всегда 0,7 http://habshuncoy.webcindario.com/caviho/rayonniy-monitoring-po-geometrii-9-klass. php2019-11-04всегда0.5 http://habshuncoy.webcindario.com/caviho/gdz-po-konturnim-kartam-po-geografii-7-klass-drofa.php2019-11-06 всегда0,8 http://habshuncoy.webcindario.com/caviho/atlas- po-geografii-5-klass-drofa.php2019-11-08 всегда0.5 http://habshuncoy.webcindario.com/caviho/rabochaya-tetral-4-klass-kuzovlev.php2019-11-10 всегда0.7http: // habshuncoy. webcindario.com/caviho/skachat-otveti-po-literaturnomu-chteniyu-3-klass-boykina.php2019-11-13 всегда0,5 http://habshuncoy.webcindario.com/caviho/testovie-zadaniya-po-obshestvoznaniyu-10 класс-1-полугодие.php2019-11-16всегда0,6 http://habshuncoy.webcindario.com/caviho/didakticheskie-materiali-po-himii-8-klass-gorshkova-s-temami.php2019-11-18 всегда0,6 http://habshuncoy.webcindario. com / caviho / gdz-po-leteraturnomu-chteniyu-4-klass-buneev.php2019-11-21always0.6http: //habshuncoy.webcindario.com/caviho/skachat-gdz-po-nemetskomu-za-10-11- klass.php2019-11-22всегда0.6 http://habshuncoy.webcindario.com/caviho/gdz-po-angliskomu-4-klass-znnikitenko.php2019-11-23всегда0.7http: //habshuncoy.webcindario/caviho.com/caviho тетрад-для-практических-работ-по-географии-9-класс-аг-стадник.php2019-11-24 всегда0,5 http://habshuncoy.webcindario.com/caviho/uchebniki-dlya-7-klassa-na-planshet.php2019-11-26 всегда0,7 http://habshuncoy.webcindario.com/caviho/dlya- 6-го-класс-сканворд-по-теме-культура-халифата-сканворд.php2019-11-28всегда0.5 http://habshuncoy.webcindario.com/caviho/reshenie-nomerov-po-algebre-8-klass-mordkovich. php2019-11-29 всегда0.5 http://habshuncoy.webcindario.com/caviho/spotlight-5-klass-workbook-audio.php2019-11-30 всегда0.6 http://habshuncoy.webcindario.com/caviho/spishi-ru- биология-9-класс.php2019-12-02 всегда0.6 http://habshuncoy.webcindario.com/caviho/gdz-po-rabochey-tetradi-pol-informatike-klass.php2019-12-03 всегда0.5 http://habshuncoy.webcindario.com/caviho/ решебник-по-литературному-чтению-2-класс-м-бойкина.php2019-12-04всегда0.7 http://habshuncoy.webcindario.com/caviho/radetskiy-didakticheskiy-material-gdz-10-klass-19-skachat.php 12-05всегда0.7 http://habshuncoy.webcindario.com/caviho/gdz-algebra-i-nachalo-matematicheskogo-analiza-10-klass-mordkovich-profilniy-uroven.php2019-12-06 всегда0.6 http://habshuncoy.webcindario.com/caviho/gdz-po-istorii-klass-aa-danilov.php2019-12-08 всегда0,6 http://habshuncoy.webcindario.com/caviho/students-book-9-klass- 16-izdanie.php2019-12-09 всегда0,6 http://habshuncoy.webcindario.com/caviho/rabochaya-tetrad-kaufman-7-klass.php2019-12-10 всегда0,6 http://habshuncoy.webcindario.com/caviho/ gdz-gotovie-domashnie-zadaniya-po-geometrii-za-klass.php2019-12-12 всегда 0.8 http://habshuncoy.webcindario.com/caviho/7-klass-gdz-po-algebre-mordkovich-i-dr. php2019-12-13всегда0.5 http://habshuncoy.webcindario.com/caviho/knigi-10-klass-skachat-na-planshet.php2019-12-14всегда0,8 http://habshuncoy.webcindario.com/caviho/plani-konspekti-urokov-po- obzh-9-klass.php2019-12-16 всегда0.8 http://habshuncoy.webcindario.com/caviho/skachat-testi-po-vseobshey-istorii-7-klass.php2019-12-16 всегда0.7 http://habshuncoy. webcindario.com/caviho/tetrad-dlya-laboratornih-rabot-po-biologii-7-klass-otveti-poperenko.php2019-12-17 всегда0,7 http://habshuncoy.webcindario.com/caviho/istoriya-5-klassa- все-ответы-в-тетради.php2019-12-18всегда0.6 http://habshuncoy.webcindario.com/caviho/gdz-po-gia-9-klass-pod-redaktsiey-fflisenko-syukulabuhova-2020g.php2019-12-19 всегда0.7http: // habshuncoy. webcindario.com/caviho/reshebnik-po-himii-10-klass-profilniy-uroven.php2019-12-20всегда0.8 http://habshuncoy.webcindario.com/caviho/gdz-po-angliyskumu-8-klass-pismo- drugu.php2019-12-22всегда0.6http: //habshuncoy.webcindario.com/caviho/reshebnik-umniki-i-umnitsi-3-klass.php2019-12-23всегда0.8http: //habshuncoy.webcindario.com/hytepur/ always1http: // habshuncoy.webcindario.com/hyhytepur/gdz-po-pechatnoy-tetradi-biologiya-7.php2019-10-25всегда0.7 http://habshuncoy.webcindario.com/hyhytepur/bzhd-uchebnik.php2019-10-27 всегда0.6http:// habshuncoy.webcindario.com/hyhytepur/skachat-uchebnik-8-klliteraturazvinyatskovskiy-filenko.php2019-10-29 всегда 0,6http://habshuncoy.webcindario.com/hyhytepur/aprimkevich-zbornik-gizadach php2019-10-31 всегда0,6 http://habshuncoy.webcindario.com/hyhytepur/biologiya-10-11-gdz.php2019-11-02 всегда0,8 http://habshuncoy.webcindario.com/hyhytepur/test-po-biologii-7-tip-chervi-proverka-svoih-znaniy.php2019-11-04 всегда0.6 http://habshuncoy.webcindario.com/hyhytepur/bendrikov-fizika-gdz-reshebnik- skachat.php2019-11-06 всегда0,7 http://habshuncoy.webcindario.com/hyhytepur/gdz-po-uchebniku-lg-peterson-gv-dorofeev.php2019-11-08 всегда0,8 http://habshuncoy.webcindario.com/ hyhytepur / gdz-po-algebre-alimov.php2019-11-10 всегда0.5 http://habshuncoy.webcindario.com/hyhytepur/urovni-srednevekovoy-kulturi-tablitsa.php2019-11-13 всегда0.5 http://habshuncoy.webcindario.com/hyhytepur/pnsh3klass-kontrolnaya-rabota-za-2-yu-chetvert.php2019-11-16 всегда0,5 http://habshuncoy.webcindario.com/hyhytepur/uchebniki-po-t pravu-skachat-2020-god-format-doc.php2019-11-19 всегда0.6 http://habshuncoy.webcindario.com/jywekux/always1http: //habshuncoy.webcindario.com/jywekux/nem-yaz-5-klas- rabochaya-tetrad-2020.php2019-10-25 всегда0,7 http://habshuncoy.webcindario.com/jywekux/gotov-zavdannya-z-konturno-karti-10-klas.php2019-10-27всегда0,5 http://habshuncoy.webcindario.com/jywekux/ya-hotel-chtob-v-serdtse-spominalo-sad-i-leto-gdz.php2019-10-29 всегда0,6 http://habshuncoy.webcindario.com/jywekux/obshaya-elektrotehnika-s- osnovami-elektroniki-otveti.php2019-10-31 всегда0.6 http://habshuncoy.webcindario.com/jywekux/gdz-evgladchenko-gakasyan-mapoznyak-diholin.php2019-11-02 всегда0.8http: //habshuncoy. jywekux / gdz-6-klas-ros-mova-balandina-degtereva-lebedenko.php2019-11-04 всегда0.8 http://habshuncoy.webcindario.com/jywekux/dudnitsi-yu-p-geometriya-gdz-kla.php2019-11-06 всегда0,6 http://habshuncoy.webcindario.com/jywekux/gotovie-domashnie-zadaniya-k-uchebniku-istorii.php2019-11-09 всегда0,5 http://habshuncoy.webcindario.com/jyve-kux/ot k-labratornoy-rabote-po-fizike-ilechenko-kulikovskiy-7klass.php2019-11-10всегда0.6 http://habshuncoy.webcindario.com/jywekux/gdz-6-klas-zagalna-geografya-oyaskuratovich.php2019-11-10 13всегда0,8 http://habshuncoy.webcindario.com/jywekux/latin-reshebnik-gorodkova.php2019-11-16 всегда0,6 http://habshuncoy.webcindario.com/jywekux/gdz-k-uchebniku-alimova-10-11- клас.php2019-11-19 всегда0,5 http://habshuncoy.webcindario.com/cikom/always1 http://habshuncoy.webcindario.com/cikom/gdz-7-klass-angliyskiy-nataliya-koltko-bogdan.php2019-10-26. 6 http://habshuncoy.webcindario.com/cikom/rabochaya-tetrad-po-geografii-8-klass-kim-otveti-slayd-shou-video.php2019-10-27всегда0,6http: //habshuncoy.webcindario.com/ cikom / informatika-7-klass-semakin-gdz.php2019-10-29always0.7 http://habshuncoy.webcindario.com/cikom/istoriya-za-7-klass-danilov-chitat.php2019-10-31always0.7http: // habshuncoy.webcindario.com/cikom/stranichki-lyuboznatelnih-moro-3-klass-igra-odinnadtsat-palochek.php2019-11-02 всегда0.7 http://habshuncoy.webcindario.com/cikom/pochva-vazhneyshaya-chast-ekosistemi-pre 3-klass.php2019-11-04 всегда0.8 http://habshuncoy.webcindario.com/cikom/po-uchebniku-fiziki-8-klassa-asperishkin-testi-ik-nemu-otveti.php2019-11-06 всегда0.8http: //habshuncoy.webcindario.com/cikom/gdz-klass-po-anglisskomu-yaziku.php2019-11-09 всегда0.5 http://habshuncoy.webcindario.com/cikom/konspekt-zadacha-nbistomina-2-klass.php2019-11-10 всегда 0,6 http://habshuncoy.webcindario.com/cikom/angliyskiy-5-klass-oksana-karpyuk-otveti-k-rabochey-tetradi.php2019-11-13 всегда 0,6 http://habshuncoy.webcindario. com / cikom / pomogite-perevesti-tekst-s-angliyskogo-na-russkiy-5-klass.php2019-11-16always0.6http: //habshuncoy.webcindario.com/cikom/istoriya-10-klass-saharov-1- chast-gdz.php2019-11-19 всегда0,8 http://habshuncoy.webcindario.com/cikom/otveti-na-krossvordi-vedyushkin-6-klass.php2019-11-21 всегда0,8 http://habshuncoy.webcindario.com/ циком / презентация-история-владимирского-край-класс.php2019-11-22всегда0,7 http://habshuncoy.webcindario.com/cikom/bilyanina-10-klass-gdz.php2019-11-23всегда0,8 http://habshuncoy.webcindario.com/cikom/gdz-po-geografii- k-paragrafam-10-klass-maksakovskiy.php2019-11-25 всегда0,5 http://habshuncoy.webcindario.com/cikom/gdz-po-geografii-klass-po-gerasimovoy-i-neklyukovoy.php2019-11-26 всегда0. 7 http://habshuncoy.webcindario.com/cikom/gdz-po-angliyskomu-11-klass-new-millenium-english.php2019-11-28 всегда 0,7 http://habshuncoy.webcindario.com/cikom/gdz-klass- 2020-года-випуска.php2019-11-29всегда0.6 http://habshuncoy.webcindario.com/cikom/gdz-po-obshestvoznaniyu-11-klass-ln-bogolyubova.php2019-12-01 всегда0.6 http://habshuncoy.webcindario.com/cikom краткий-консект-по-истории-россии-8-класс-данилов-параграфа-5.php2019-12-02всегда0.7 http://habshuncoy.webcindario.com/cikom/gdz-po-biologii-6-klass-paragraf- 18-тпо-пасечник-снисаренко-2020-onlain.php2019-12-03всегда0.7 http://habshuncoy.webcindario.com/cikom/olimpiada-po-obshestvu-za-7-klass-2020-god.php2019-12- 04всегда0,5 http://habshuncoy.webcindario.com/cikom/reshebnik-istoriya-drevnego-mira-rabochaya-tetrad-5-klass-1-chast-goder-zadanie.php2019-12-05 всегда0.7 http://habshuncoy.webcindario.com/cikom/reshenie- Упражнения-426-по-учебнику-6-класс-сильвова.php2019-12-06всегда0.6 http://habshuncoy.webcindario.com/cikom/gdz-po-angliyskomu-5-klass-kuzovlev-lapa-duvanova-kostina- kuznetsova.php2019-12-08 всегда0,6 http://habshuncoy.webcindario.com/cikom/gdz-po-angliyskomu-8-klass-mz-biboletova.php2019-12-09 всегда0,7 http://habshuncoy.webcindario.com / cikom / 7-класс-новая-история-8-9-параграф-художники-i-ih-proizvedeniya-baranov.php2019-12-10всегда0.8http: //habshuncoy.webcindario.com/cikom/uroki-angliyskogo- 3-класс-задание-24-номер-6-video.php2019-12-12всегда0.6 http://habshuncoy.webcindario.com/cikom/kompleksnaya-kontrolnaya-rabota-i-polugodie-2-klass-fgos.php2019- 12-14 всегда0,5 http://habshuncoy.webcindario.com/cikom/raznourovnevie-zadaniya-po-teme-reshenie-lineynih-neravenstv-9-klass.php2019-12-15 всегда0,7 http://habshuncoy.webcindario.com/ cиком / решебник-по-англ-яз-5-класс-вербицкая.php2019-12-16 всегда 0,8 http://habshuncoy.webcindario.com/cikom/perevod-teksta-s-angliyskogo-na-russkiy-8-klass-kaufman.php2019-12-16 всегда 0,8 http://habshuncoy.webcindario. com / cikom / konspekt-uroka-vostok-v-nachale-20-veka-9-klass-noveyshaya-istoriya.php2019-12-17 всегда0.5 http://habshuncoy.webcindario.com/cikom/rabota-so-slovaryami- vo-2-klasse-pnsh.php2019-12-18 всегда0,8 http://habshuncoy.webcindario.com/cikom/gdz-po-uchebniku-geografii-klass.php2019-12-19 всегда0,6 http://habshuncoy.webcindario. ком / циком / плани-по-биологии-7-класс-латюшин.php2019-12-20всегда0,8 http://habshuncoy.webcindario.com/cikom/galitskiy-reshebnik-8-klass-21.php2019-12-22всегда0,8 http://habshuncoy.webcindario.com/cikom/skachat-gdz- по-истории-9-класс-загладин-всеобщая-история.php2019-12-23always0.5 http://habshuncoy.webcindario.com/cikom/perevod-teksta-po-angliyskomu-5-klass.php2019-12-24always0. 5 http://habshuncoy.webcindario.com/vuzoqozaw/always1http: //habshuncoy.webcindario.com/vuzoqozaw/gdz-algebra-klass-yun-makarichev-ng-mindyuk-ki-neshkov.php2019-10-26.5 http://habshuncoy.webcindario.com/vuzoqozaw/gdz-po-fizike-k-uchebniku-gladkova-i-tsodikova.php2019-10-27всегда0,8http: //habshuncoy.webcindario.com/vuzoqozaw-po/reshebniku prirodoznavstvo-5-rjnbr.php2019-10-29 всегда0.5 http://habshuncoy.webcindario.com/vuzoqozaw/gdz-po-algebre-8-klass-makarichev-pod-redaktsiey-telyakovskogo-17e-izdanie. 31всегда0,6 http://habshuncoy.webcindario.com/vuzoqozaw/gdz-sbornik-testov-ege-po-algebre.php2019-11-02 всегда0,7 http://habshuncoy.webcindario.com/vuzoqozaw/gdz-po-algedre классов-макаричев-14-издание.php2019-11-04всегда0.6 http://habshuncoy.webcindario.com/vuzoqozaw/reshebnik-po-algebre-9-klass-makarichev-2020-smotret.php2019-11-06 всегда0.6 http://habshuncoy.webcindario.com/ vuzoqozaw / search-gdz-po-program-razumovskoy.php2019-11-09 всегда0.5 http://habshuncoy.webcindario.com/vuzoqozaw/otveti-na-testi-po-fizike-9klass-gromtseva.php2019-11-10 всегда0. 5 http://habshuncoy.webcindario.com/vuzoqozaw/konspekt-uroka-po-sbo-na-temu-uhod-za-golovnimi-uborami.php2019-11-14 всегда0,6 http://habshuncoy.webcindario.com / vuzoqozaw / skachat-bez-registratsii-po-fgos-dlya-zabotlivih-roditeley-all-domashnie-zadaniya-2klassreshebnik-iv.php2019-11-16always0.7
8http: //habshuncoy.com / sykixox / srezi-po-algebre-11-klass-2020.php2019-11-09 всегда0.6 http://habshuncoy.webcindario.com/sykixox/gdz-11-klass-algebra-bevz-bevz-vladimirova.php2019- 11-11всегда0,7 http://habshuncoy.webcindario.com/sykixox/aalobzhanidze-tetrad-trenazher-po-geografii-5-6-klass-skachat.php2019-11-14 всегда0,6 http://habshuncoy.webcindario.com/ sykixox / podgotovka-k-olimpiade-po-biologii-9-klass.php2019-11-17всегда 0,7 http://habshuncoy.webcindario.com/sykixox/kak-otvetit-na-vopros-10-po-istorii-6- класс-на-странице-152-по-авторам-ев-агибалова-игм-донской.php2019-11-19 всегда0.5 http://habshuncoy.webcindario.com/sykixox/gdz-algebra-i-nachala-matematicheskogo-analiza-10-klass-kolyagin-reshebnik-onlayn.php2019-11-21 всегда0.8http: // habshuncoy.webcindario.com/sykixox/olimpiada-po-biologii-7-9-klass.php2019-11-22 всегда0,8 http://habshuncoy.webcindario.com/sykixox/otveti-po-tetradi-literaturnoe-chtenie-2- klass-buneev.php2019-11-23всегда0.7 http://habshuncoy.webcindario.com/sykixox/gdz-po-algebre-pod-redaktsiey-satelyakovskogo-9-klassa.php2019-11-25 всегда0.7http: // habshuncoy.webcindario.com/sykixox/gdz-ladizhenskaya-9-klass.php2019-11-26всегда 0,7 http://habshuncoy.webcindario.com/sykixox/okruzhayushiy-mir-3-klass-rabochaya-tetrad-pleshakov-1-chast- kartinki-str11.php2019-11-28всегда0.7 http://habshuncoy.webcindario.com/sykixox/algebra-na-kazahskom-yazike-za-9-klass-avtori-abilkasimova-bekboev-reshenie-zadachi-nomer-97. php2019-11-29 всегда0,8 http://habshuncoy.webcindario.com/sykixox/algebra-8-klass-merzlyak-s-uglublennim-izucheniem-reshebnik-onlayn.php2019-12-01 всегда0,7 http://habshuncoy.webcindario.com/sykixox/gdz-po-algebre-klass-avtori-agmordkovich-laaleksandrova-pdf-skachat.php2019-12-02 всегда0,6 http://habshuncoy.webcindario.com/sykixox/obzh-6-klass-rabochaya- tetrad-zadanie-4.php2019-12-03 всегда0,8 http://habshuncoy.webcindario.com/sykixox/gotovie-resheniya-po-algebre-8-klass-telyakovskiy.php2019-12-04 всегда0,6 http://habshuncoy. webcindario.com/sykixox/gdz-po-rabochey-tetradi-kotova-7-klass.php2019-12-05 всегда0.8 http://habshuncoy.webcindario.com/sykixox/reshebnik-po-rabochey-tetradi-4-klassa- наслаждайся английским.php2019-12-06всегда0,6 com / sykixox / ulp-po-vseobshey-istorii-srednih-vekov-rabochaya-tetrad-6-klass.php2019-12-09 всегда0.5 http://habshuncoy.webcindario.com/sykixox/reshenie-algebri-za-9- klass-avtor-shinibekov.php2019-12-10 всегда 0.8 http://habshuncoy.webcindario.com/sykixox/gdz-po-istorii-6-klass-rabochaya-tetrad-kryuchkova-k-paragrafu.php2019-12-12 всегда0.6 http://habshuncoy.webcindario.com/sykixox/uslovie-k-zadache-pro-truboprovodi-3-klass.php2019-12-14всегда0,6http: //habshuncoy.webcindario.com/sykixox/gotoviy-kompleks-kompleks-niy-aniy- teksta-9-klass-6-tekst.php2019-12-15всегда0.8 http://habshuncoy.webcindario.com/sykixox/gdz-po-istorii-7-klass-yudovskaya-konspekt-25-paragrafa.php2019-12- 16 всегда0,6 http://habshuncoy.webcindario.com/sykixox/y-eshe-6-klass.php2019-12-16всегда0,5 http://habshuncoy.webcindario.com/sykixox/gdz-po-fizike-10-klass- 2020g.php2019-12-17всегда0.5 http://habshuncoy.webcindario.com/sykixox/gdz-po-biologii-chelovek-uchebnik-dlya-8-klassa-obsheobrazovatelnoy-shkoli.php2019-12-18always0.8http: //habshixuncoy.webcindario.com/sykixoxy.webcindario.com/sykix diktanti-po-teme-leksika-5-klass.php2019-12-19 всегда0.6 http://habshuncoy.webcindario.com/sykixox/gdz-po-algebre-dlya-10-11-klassov-alimov-sha-kolyagin- yum-izdatelstvo-prosveshenie-ghjdthm-ct-z.php2019-12-21always0.8 http://habshuncoy.webcindario.com/sykixox/skachat-uchebnik-po-biologii-10-11-klass.php2019-12-22always0.7 http://habshuncoy.webcindario.com/cimyrire/always1http: //habshuncoy.webcindario.com/cimyrire/zadacha-4-po-matematike-3-klass-str-4-demidova-kozlova-tonkih19-chast.php20 10-24всегда0,5 http://habshuncoy.webcindario.com/cimyrire/gdz-po-matematike-3-klass-istomina-593-zadacha.php2019-10-26всегда0,5 http://habshuncoy.webcindario.com/cimyrire/ конспект-открытого-урока-математики-школа-россии-в-1-класс-с-самоанализом.php2019-10-28всегда0.5http: //habshuncoy.webcindario.com/cimyrire/uchebnik-3-klassa-po-mate часть-1-тонких-демидова-с-9-упр-7.php2019-10-30всегда0.8 http://habshuncoy.webcindario.com/cimyrire/matematika-domashnyaya-rabota-gdz-4-klass.php2019-11-01 всегда0.6 http://habshuncoy.webcindario.com/cimyrire/vopros с-ответами-для-тестирования-учителей-начальных-классов.php2019-11-03всегда0.5 http://habshuncoy.webcindario.com/cimyrire/matematika-4-klass-chast-1-iz-2-mi-moro- ma-bantova-gv-beltyukova-i-dr-2020-reshebnik.php2019-11-05 всегда0.6 http://habshuncoy.webcindario.com/cimyrire/razrabotka-laboratornoy-raboti-po-biologii-8-klass-opredelenie- норма-рационального-питания.php2019-11-07всегда0,7 http://habshuncoy.webcindario.com/cimyrire/otveti-np-olimpialu-po-obshestvoznaniyu-8-klass-2020.php2019-11-09 всегда0,7 http://habshuncoy.webcindario.com/ cimyrire / lgstadnik-matematika-protochniy-zoshit-6-klass.php2019-11-11всегда0.7 http://habshuncoy.webcindario.com/cimyrire/dz-4-klass-po-matematike-moro-nomer.php2019-11- 14всегда0,6http: //habshuncoy.webcindario.com/cimyrire/gotovie-domashnie-zadaniya-5-klass-matematika-kozlova-rubin.php2019-11-17 всегда0,7 http://habshuncoy.webcindario.com / cimyrire / reshebnik-po-matematike-4-klass-iiarginskaya-eiivanovskaya-snkormishina.php2019-11-19 всегда0.5http: //habshuncoy.webcindario.com/cimyrire/vi-sirotin-9-klass-gdz-gdz 11-21всегда0,7 http://habshuncoy.webcindario.com/cimyrire/biologiya-cheloveka-8-klass-kolesov-18-paragraf.php2019-11-22 всегда0,5 http://habshuncoy.webcindario.com/cimyrire/gdz- praktikum-10-11-klass-po-ekonomsike.php2019-11-24 всегда0.6 http://habshuncoy.webcindario.com/cimyrire/gdz-7-klass-angl-yaz-spotlight.php2019-11-25 всегда0.5 http://habshuncoy.webcindario.com/cimyrire/rabochaya-programma-po-matematike-3-klass-davidov-gorbov.php2019-11-27 всегда0,5 http://habshuncoy.webcindario.com/cimyripore-/gdz geometrii-9-klass-atanasyan-2004-god-zo.php2019-11-29always0.7 http://habshuncoy.webcindario.com/cimyrire/gdz-informatsiya-vokrug-nas-5-klass-binom.php2019-11- 29всегда0,7 http://habshuncoy.webcindario.com/cimyrire/proekti-dlya-tehnologiy-9-klass.php2019-12-01 всегда0,8 http://habshuncoy.webcindario.com/cimyrire/otveti-po-rabochey-tetradki-tetradki всемирной-истории-11-класс.php2019-12-02 всегда0,5 http://habshuncoy.webcindario.com/cimyrire/matematika-2-klass-elkonina-davidova.php2019-12-03 всегда0,5 http://habshuncoy.webcindario.com/cimyrire/otveti-po matematike-5-klass-peterson-1-chast-2020.php2019-12-04всегда0.5 http://habshuncoy.webcindario.com/cimyrire/gdz-po-matem-klass-vilenkin-zhohov-2020god.php2019-12- 05 всегда0,8 http://habshuncoy.webcindario.com/cimyrire/zadacha-9-urok-58-matematika-geydman-3-klass.php2019-12-06 всегда0,6 http://habshuncoy.webcindario.com/cimyrire/kdr- 9-класс-математика-апрел.php2019-12-08 всегда0,6 http://habshuncoy.webcindario.com/cimyrire/zadachi-dlya-2-klassa-2-chetvert-matematika.php2019-12-09 всегда0,6 http://habshuncoy.webcindario.com/cimyrire/ математика-6-класс-контрольно-измерительные-материалы-решебник-lp-popova.php2019-12-10 всегда0,8 http://habshuncoy.webcindario.com/cimyrire/gdz-po-matematike-5-klass-2019-gambarin.php 12-13всегда0,8 http://habshuncoy.webcindario.com/cimyrire/kontrolnie-raboti-po-matematike-peterson-2-klass-1-chetvert.php2019-12-14 всегда0,8 http://habshuncoy.webcindario.com/cimyrire/kontrolnaya-rabota-po-matematike-3-klass-za-1-polugodie-po-moro.php2019-12-15 всегда0,6 http://habshuncoy.webcindario.com/cimyrire/kontrolnie-raboti- по-математике-и-русскому-языку-за-1-полугодие-в-4-класс-школа-россии.php2019-12-16всегда0.7 http://habshuncoy.webcindario.com/cimyrire/matematika-srez-10- klass-otveti-2-variant.php2019-12-16всегда0.5 http://habshuncoy.webcindario.com/cimyrire/gotovie-domashnie-zadaniya-11-klass-rus-yaz-rozental.php2019-12-17 всегда0.6http: // habshuncoy.webcindario.com/cimyrire/gotovaya-konturnaya-karta-8-klass.php2019-12-18всегда 0,7 http://habshuncoy.webcindario.com/cimyrire/kalendarno-tematicheskoe-planirovanie-po-matematike-chteniyu-2-klass fgos-shkola-rossii.php2019-12-20всегда0.7 http://habshuncoy.webcindario.com/cimyrire/itogovaya-kontrolnaya-rabota-po-vseobshey-istorii-8-klass.php2019-12-21 всегда0.7http: // habshuncoy.webcindario.com/cimyrire/gdz-po-istorii-9-klass-rabochaya-tetrat-chast-pervaya.php2019-12-22always0.6http://habshuncoy.webcindario.com / cimyrire / poisk-skachat-gotovie-domashnie-zadaniya-po-matematike-za-klass.php2019-12-23always0.6 http://habshuncoy.webcindario.com/wuzam/always1http: //habshuncoy.com/webcuncoy.com wuzam / konspekt-uroka-po-fgos-po-russkomu-yaziku.php2019-10-25всегда0.6http: //habshuncoy.webcindario.com/wuzam/uchebnik-po-angliyskomu-yaziku-5-klass-bibol-tova-chit 2020-god.php2019-10-26всегда0,8 http://habshuncoy.webcindario.com/wuzam/angliyskiy-dlya-psihologov-kovalenko-reshebnik.php2019-10-28всегда0,7 http://habshuncoy.webcindario.com/wuzam/gdz-po-angliyskomu-yaziku-5-klass-progress-chek.php2019-10-30 всегда0.6 http://habshuncoy.webcindario.com/wuzam/gotovie-domashnie-zadaniya-2-klass- angliyskiy-yazik-nesvit.php2019-11-01 всегда 0,6 http://habshuncoy.webcindario.com/wuzam/reshebnik-po-belorusskomu-yaziku-4-klass-tm-valinets-ie-ratnikava-is-rouda-pp- shuba-1-ch-2020-g.php2019-11-03 всегда0.7 http://habshuncoy.webcindario.com/wuzam/gdz-angliyskiy-yazik-8klass-kuzovlev-vp.php2019-11-05 всегда0.5 http://habshuncoy.webcindario.com/wuzam/gdz-angliyskiy-yazik-8klass-kuzovlev-vp.php2019-11-05 всегда0.5 habshuncoy.webcindario.com / wuzam / angliyskiy-yazik-uchebnik-dlya-10-11-klassov-gdz.php2019-11-07 всегда0.5 http://habshuncoy.webcindario.com/wuzam/gdz-po-angliyskomu-yaziku-5-klass- perevod-ab.php2019-11-09 всегда0.5 http://habshuncoy.webcindario.com/wuzam/reshebnik-po-tatarskomu-yaziku-5-klass-16-gimnaziya.php2019-11-11 всегда0.7http: // habshuncoy. webcindario.com/wuzam/gdz-po-angliyskomu-yaziku-8-klass-nesvit-zadanie-4-stranitsa-13.php2019-11-15 всегда0.8 http://habshuncoy.webcindario.com/wuzam/gdzpo-nemetskomu- yaziku-klass.php2019-11-17всегда0.8 http://habshuncoy.webcindario.com/wuzam/reshebnik-angliyskiy-yazik-rabochaya-tetrad-myasoedova-4klass-gde-skachat.php2019-11-20 всегда0,8 http://habshuncoy.webcindario.com/zwuz по-анлискаму-языку-за-классавтор-афанасьева-и-верешагина.php2019-11-21всегда0.6http: //habshuncoy.webcindario.com/wuzam/perevodi-tekstov-kaufman-11-klass-angliyskiy-yaz 11-22всегда0.7 http://habshuncoy.webcindario.com/wuzam/nemetskiya-yazik-rabochaya-tetrad-7-klass-gdz-bim-sadomova-fomicheva.php2019-11-24 всегда0.5 http://habshuncoy.webcindario.com/wuzam/reshebnik-po-angliyskomu-yaziku-7-klass-matriks.php2019-11-25всегда0.8http: //habshuncoy.webcindario.com/wuzam/dz-angliyskiy-yazik 11-yuhnel.php2019-11-27 всегда0.5 http://habshuncoy.webcindario.com/wuzam/gdz-tatarskiy-yazik-6-klass-spisat.php2019-11-29всегда0,8 http://habshuncoy.webcindario.com/ wuzam / сайт-гарфилда-готовые-домашние-задания-по-английскому-yaziku.php2019-11-29всегда0.8 http://habshuncoy.webcindario.com/wuzam/7-klassaudiozapis-angliyskogo12.php20.7 http://habshuncoy.webcindario.com/wuzam/uchebnik-po-angliyskomu-yaziku-mechel-harris-irina-sokolova.php2019-12-02 всегда0,5 http://habshuncoy.webcindario.com/wuzam/gdz-po английскому-языку-10-класс-биболетова-рабочая-тетрад-1-onlayn.php2019-12-03 всегда0,8 http://habshuncoy.webcindario.com/wuzam/tekst-po-angliyskomu-yaziku-7-klass-spot. php2019-12-04всегда0.8 http://habshuncoy.webcindario.com/wuzam/angliyskiy-yazik-3-klass-rabochaya-tetrad-otveti-bikova-2020.php2019-12-05 всегда0.7http: // habshuncoy.webcindario.com/wuzam/gdz-frantsuzskiy-yazik-grigoreva-gorbacheva-10-11-klass.php2019-12-06 всегда0.5 http://habshuncoy.webcindario.com/wuzam/besplatnie-otveti-po-tatarskomu-yaziku- ksfethullova-gdz-6-klass.php2019-12-09 всегда 0,8 http://habshuncoy.webcindario.com/wuzam/chitat-uchebnik-po-angliyskomu-yaziku-kuzovlev-10-klass-onlayn.php2019-12-09 всегда. 8 http://habshuncoy.webcindario.com/wuzam/tekst-po-angliyskomu-yaziku-alla-nesvit.php2019-12-11всегда0,6http: //habshuncoy.webcindario.com/wuzam/reshebnik-po-angliyskomu-yaziku-yaz 7-класс-ваулина-учебник.php2019-12-13всегда 0,7 http://habshuncoy.webcindario.com/tolyvoxor/always1 http://habshuncoy.webcindario.com/tolyvoxor/biologiya-9-ponomareva-samostoyatelnie.php2019-10-25всегда //0,8http: webcindario.com/tolyvoxor/megaresheba-prakticheskaya-po-biologii.php2019-10-26всегда 0,7 http://habshuncoy.webcindario.com/tolyvoxor/reshebnik-kontrolnih-rabot-5-6-matematika-kuznetsova-20mina 10-29 всегда 0,7 http://habshuncoy.webcindario.com/tolyvoxor/praktikum-po-visshey-matematike-dlya-ekonomistov-kremer-gdz.php2019-10-30всегда0,6 http://habshuncoy.webcindario.com/tolyvoxor/gdz-rabochaya-tetrad-po-matematike-potapov-i-shevkin.php2019-11-01 всегда0,6 http://habshuncoy.webcindario.com/ tolyvoxor / gotovie-domashnie-zadaniya-po-matematike-n-ya-vilennkin.php2019-11-03 всегда0.5 http://habshuncoy.webcindario.com/tolyvoxor/gdz-po-obshage.php2019-11-06 всегда: 0.6httpalways //habshuncoy.webcindario.com/tolyvoxor/hachaturyan-uchebnik-investitsii-skachat.php2019-11-07 всегда0.8 http://habshuncoy.webcindario.com/tolyvoxor/vdpovd-na-zavdannya-matematika-boglog.php2019-11-09всегда0,7 http://habshuncoy.webcindario.com/tolyvoxor/gdz-galitskiy.php2019-11-11всегда0,6 http://habshuncoy.webcindario.com/tolyvoxor/gdz-po-muzike-tvorcheskaya-tedorcheskaya. php2019-11-15 всегда0,5 http://habshuncoy.webcindario.com/tolyvoxor/ege-12-matematika-etap-1-klass-11-variant-2-otveti.php2019-11-17 всегда0,6 http://habshuncoy. webcindario.com/tolyvoxor/gdz-dlya-6-go-klassavilenkin-nya-chesnokov-as-i-drpo-matematike.php2019-11-20всегда0,5 http://habshuncoy.webcindario.com/tolyvoxor/zanimatelnie-zadaniya- по-математике-для-начальной-школы.php2019-11-21 всегда0,5 http://habshuncoy.webcindario.com/tolyvoxor/kontrolnaya-po-matematiki-vilenkin.php2019-11-22всегда0,8 http://habshuncoy.webcindario.com/kikilahub/always1httpuncohub/always1http: webcindario.com/kikilahub/chasti-pochek-rasteniy-6-klass-pobeg-i-pochki.php2019-10-25всегда0,5 http://habshuncoy.webcindario.com/kikilahub/algebra-7-klass-testi-s- otvetami-onlayn.php2019-10-27 всегда0,7 http://habshuncoy.webcindario.com/kikilahub/olimpiada-po-fizike-8-klass-2020-god-zadaniya-i-otveti.php2019-10-29 всегда0.6 http://habshuncoy.webcindario.com/kikilahub/chitat-uchebnik-biboletova-7-klass.php2019-10-30 всегда0,6 http://habshuncoy.webcindario.com/kikilahub/7-klass-onlayn-uchebnik- biologii-zaharov-vb.php2019-11-01 всегда0.6 http://habshuncoy.webcindario.com/kikilahub/uchebnik-4-klassa-po-russkomu-yaziku-vinogradovo-onlayn.php2019-11-03 всегда0.7http: // habshuncoy.webcindario.com/kikilahub/reshebnik-po-geografii-za-7-klass-po-pechatnoy-tetradi-sirotin-bez-registratsii.php2019-11-06 всегда0.6http://habshuncoy.webcindario.com/kikilahub/gdz-rabochaya-tetrad-po-biologii-11-klass-ponomareva-onlayn.php2019-11-07 всегда0.7 http://habshuncoy.webcindario.com/kikilahub/rabochaya-tetrad-po-obshestvu- gdz-onlayn-7-klass-katova.php2019-11-10всегда0.6 http://habshuncoy.webcindario.com/kikilahub/gotovie-domashnie-zadaniya-po-literature-klass-po-uchebniku-maratsmana.php2019-11-10 12всегда0,8 http://habshuncoy.webcindario.com/kikilahub/kontrolniy-diktant-3-klass-zankov-nechaeva.php2019-11-15 всегда0,6 http://habshuncoy.webcindario.com / kikilahub / otveti-na-prakticheskuyu-rabotu-5-po-geografii-9-klass.php2019-11-17всегда0.5http: //habshuncoy.webcindario.com/kikilahub/reshenie-zadach-po-matematike-4- klass-bogdanovich-mv-onlayn-nomer-809.php2019-11-20всегда0.7 http://habshuncoy.webcindario.com/kikilahub/gdz-po-mhk-5-klass.php2019-11-22всегда0.8http: // habshuncoy.webcindario.com/kikilahub/rabochaya-tetrad-po-literature-k-uchebniku-6-klassa-soloveva-otveti.php2019-11-22 всегда0.6http://habshuncoy.webcindario.com/kikilahub/gdz- россии-11-класс-загладин-козленко-минаков-петров.php2019-11-24 всегда 0,7 http://habshuncoy.webcindario.com/kikilahub/gotovie-domashnie-zadaniya-po-reader-kuzovleva-8-klass.php2019-11-26всегда0,6 http://habshuncoy.webcindario.com/ kikilahub / biologiya-7-klass-rabochaya-tetrad-otveti-sa-poperenko.php2019-11-27всегда0.8 klass.php2019-11-29всегда0.6 http://habshuncoy.webcindario.com/kikilahub/gdz-k-uchebniku-noveyshey-istorii-av-shubina-za-9-klass-smotret-onlayn.php2019-11-30 всегда0,7 http://habshuncoy.webcindario.com/kikilahub/reshebnik-onlayn.php2019-12-01 всегда0,6 http://habshuncoy.webcindario.com/kikilahub/zadachi-po-fizike-7-klass- f-mg.php2019-12-02 всегда 0,6 http://habshuncoy.webcindario.com/kikilahub/istoriya-tetrad-9-klass-20-veka-otveti-artasov.php2019-12-03 всегда 0,6 http://habshuncoy. webcindario.com/kikilahub/skachat-otveti-na-rabochuyu-tetrad-6-klass-erina-tm.php2019-12-05 всегда0.8 http://habshuncoy.webcindario.com/kikilahub/reshebnik-dlya-mobilnogo-telefona 7-класс-содержание.php2019-12-05 всегда0,7 http://habshuncoy.webcindario.com/kikilahub/uchebnik-himii-8-klass-rudzitis-chitat-onlayn.php2019-12-07 всегда 0,7 http://habshuncoy.webcindario.com/kikilahub андрей-игнатов-методическое-пособие-к-учебнику-ов-волобуева-и-др-россия-и-мир-10-klass.php2019-12-09всегда0.5http: //habshuncoy.webcindario.com/kikilahub/atlas- po-geografii-7-klass-smotret-avtor.php2019-12-10всегда0.7 http://habshuncoy.webcindario.com/kikilahub/gdz-po-biologiy-8-klass-rabochaya-tetrad-sonina-sapina.php2019- 12-12всегда0.7 http://habshuncoy.webcindario.com/kikilahub/razrabotki-urokov-informatiki-3-klass-goryachev.php2019-12-13всегда0.8http: //habshuncoy.webcindario.com/kikilahub/gdz-po-sbornik- по-физике-для-9-11-классов-сост-степанова-gn.php2019-12-14всегда0.6 http://habshuncoy.webcindario.com/kikilahub/gotovie-otveti-po-informatike-rabochaya-tetrad-bosova- 7-klass.php2019-12-15всегда0,8 http://habshuncoy.webcindario.com/kikilahub/gdz-biologiya-6-klass-lovyagin-onlayn.php2019-12-16всегда0,7 http://habshuncoy.webcindario.com / kikilahub / gdz-по-русскому-языку-3-класс-байкова-рабочая-тетрад-1-onlayn.php2019-12-17всегда0.8http: //habshuncoy.webcindario.com/kikilahub/onlayn-gotovie-domashnie zadaniya-po-ruskomu-yaziku-klass.php2019-12-18 всегда0.7 http://habshuncoy.webcindario.com/kikilahub/otveti-domashnee-zadanie-9-po-informatike-8-klass.php2019-12-19 всегда0. 6 http://habshuncoy.webcindario.com/kikilahub/kirik-8-klass-chitat-onlayn.php2019-12-20всегда 0,7 http://habshuncoy.webcindario.com/kikilahub/onlayn-gdz-po-biologii-9- класс-пасечник-рабочая-тетрад-задание.php2019-12-21всегда0.7 http://habshuncoy.webcindario.com/kikilahub/russkiy-yazik-8-klass-polyakova-gdz-onlayn.php2019-12-23всегда0.8 http://habshuncoy.webcindahub.com/kikilahub решебник-онлайн-5-класс-математика-с-объяснением.php2019-12-24всегда0.5 http://habshuncoy.webcindario.com/pevekeli/always1http: //habshuncoy.webcindario.com/pevekelna-i/kons podgotovke-v-shkolu.php2019-10-25always0.7 http://habshuncoy.webcindario.com/pevekeli/gdz-po-russkomu-yaziku-ege-2020-nasenina.php2019-10-27always0.8 http://habshuncoy.webcindario.com/pevekeli/gdz-russkiy-yazik-barhudarova-2002-g.php2019-10-29всегда0.5http: //habshuncoy.webcindario.com/pevekeli/sayt-nachal-httpaya-sh 1-4-prosv-ry-otveti.php2019-10-31 всегда0.7 http://habshuncoy.webcindario.com/pevekeli/reshebnik-po-russkomu-yaziku-2-na-churakova.php2019-11-01 всегда0.5http: //habshuncoy.webcindario.com/pevekeli/skachat-uchebnik-po-russkomu-yaziku-8-klass-barhudarov-torrent.php2019-11-03 всегда0,5 http://habshuncoy.webcindario.com/pevekeli/gina-dey пахнова-русский-язык.php2019-11-06 всегда0.8 http://habshuncoy.webcindario.com/pevekeli/a-vpolyakova-po-russkomu-yaziku-chast-1.php2019-11-08 всегда0.5 http://habshuncoy.webcindario.com/pevekeli/ gdz-russkiy-yazik-klass-avtor-nm-shanskiy.php2019-11-10 всегда0,5 http://habshuncoy.webcindario.com/pevekeli/gdz-pechatnaya-tetrad-po-geometrii-atanasyan-2020.php2019-11- 13всегда 0,7 http://habshuncoy.webcindario.com/pevekeli/gde-mozhno-skachat-uchebniki-dlya-plansheta.php2019-11-15всегда0,8 http://habshuncoy.webcindario.com/pevekeli/gdz-po-cher виноградов.php2019-11-18всегда0.6 http://habshuncoy.webcindario.com/pevekeli/gdz-russkiy-yazik-8-razumovskaya.php2019-11-20 всегда0.8 http://habshuncoy.webcindario.com/pevekeli/gdz-k- uchebniku-goltsova-shamshin-russkiy-yazik.php2019-11-22always0.6http: //habshuncoy.webcindario.com/pevekeli/domashnyaya-rabota-po-russkomu-yaziku-10-11-panov.php2019-11-22always0. 7 http://habshuncoy.webcindario.com/pevekeli/skachat-elektronniy-gdz-po-russkomu-yaziku-10klass-avtori-nggoltsova-ivshamshin.php2019-11-24 всегда 0,8http: // habshuncoy.webcindario.com/pevekeli/gdz-po-russkomu-socheninie-moy-dom.php2019-11-26всегда0.5 http://habshuncoy.webcindario.com/bonoju/always1http://habshuncoy.webcindario.com/bonoju/nesvit- 9-klass-uchebnik-otveti.php2019-10-25всегда0.7 http://habshuncoy.webcindario.com/bonoju/reshebniki-na-domashnie-zadaniya-2-klass-russkiy-yazik-programma-garmoniya.php2019-10- 27всегда0,7 http://habshuncoy.webcindario.com/bonoju/reshebniki-4-klassa-russkiy-yazik-buneev-buneeva.php2019-10-29всегда0,8 http://habshuncoy.webcindario.com / bonoju / spisivay-ru-5-klass-russkiy-yazik-2020-ladizhenskaya-gdz.php2019-10-31всегда0.6http: //habshuncoy.webcindario.com/bonoju/rabochaya-programma-po-russkomu-yaziku- 6-класс-ашурова-4-часа-в-неделю.php2019-11-02 всегда0.8 http://habshuncoy.webcindario.com/bonoju/gdz-russkiy-2-klass-ivanov.php2019-11-04всегда0.5http: //habshuncoy.webcindario.com/bonoju/uprazhnenie-336-6-klass-russkiy-yazik-lvova.php2019-11-06always0.7http://habshuncoy.webcindario.com/bonoju/gdz-po-russkomu-yaziku- 4-класс-вп-канакина-вг-горецкий.php2019-11-08 всегда 0,6 http://habshuncoy.webcindario.com/bonoju/kanakina-russkiy-yazik-3-klass-rabochaya-tetrad-reshebnik-stranitsa-28.php2019-11-10 всегда0,6 http://habshuncoy. webcindario.com/bonoju/gdz-reshebnik-po-russkiy-yazik-rabochaya-tetrad-dlya-2-klassa-iaisaeva.php2019-11-13всегда0.6http://habshuncoy.webcindario.com/bonoju/gdz-russkiy- yazik-6-klass-razumovskaya-4-izdanie.php2019-11-16 всегда 0,7 http://habshuncoy.webcindario.com/bonoju/skachat-russkiy-yazik-3-klass-vp-kanakina-vg-goretskiy.php2019 11-18всегда0.5 http://habshuncoy.webcindario.com/bonoju/gotovie-domashnie-zadaniya-2-klass-po-russkomu-yaziku-kalinina-chast.php2019-11-21 всегда0,5 http://habshuncoy.webcindario.com/bonoju русский-язык-4-класс-домашний-zadanie.php2019-11-22 всегда0.5 http://habshuncoy.webcindario.com/bonoju/gdz-po-russkomu-yaziku-10-klass-nikolina.php2019-11-23always0. 7 http://habshuncoy.webcindario.com/bonoju/tablitsa-po-istorii-7-klass-istoriya-novogo-vremeni-3-paragraf.php2019-11-24 всегда0,8 http://habshuncoy.webcindario.com/bonoju/ списат-домашнее-задание-по-русскому-языку-6-класс-львова-и-львов.php2019-11-26всегда0,6 http://habshuncoy.webcindario.com/bonoju/otveti-rabochaya-tetrad-5-klass-estestvoznanie-pleshakova.php2019-11-27 всегда0,5 http://habshuncoy.webcindario.com/ zadachnik-po-fizike-maron-10-klass.php2019-11-29always0.6http: //habshuncoy.webcindario.com/bonoju/gdz-russk-yaz-klass.php2019-11-30always0.5http: // habshuncoy. webcindario.com/bonoju/konspekti-urokov-po-russkomu-yaziku-4-klass-zheltovskaya.php2019-12-02 всегда0.8 http://habshuncoy.webcindario.com/bonoju/kraevaya-kontrolnaya-po-russkom-u-yaziku за-4-класс-во-2-четверти.php2019-12-02всегда0,8 http://habshuncoy.webcindario.com/bonoju/otveti-itogovaya-attestatsiya-po-russkomu-yaziku-7-klass-haustova.php2019-12-04 всегда0,7 http://habshuncoy.webcindario. com / bonoju / gotovie-domashnie-zadaniya-po-russkomu-yaziku-11-klass-vlasenkov.php2019-12-05 всегда0.8http: //habshuncoy.webcindario.com/bonoju/uprazhneniya-uchebnik-po-yazkiku-po-russkiku 5-klass-lvova.php2019-12-06 всегда0.5 http://habshuncoy.webcindario.com/bonoju/skachat-reshebnik-po-russkomu-yaziku-8-klass-lvova-lvov.php2019-12-08 всегда0.6 http://habshuncoy.webcindario.com/bonoju/gotovie-domashnie-zadaniya-russkiy-yazik-6-klass-bikova.php2019-12-09 всегда0,8 http://habshuncoy.webcindario.com/bonoju/otveti-dlya- 3-класс-по-русскому-языку-канакина.php2019-12-10всегда0.8 http://habshuncoy.webcindario.com/bonoju/reshebnik-po-tetradi-po-chteniyu-4-klass.php2019-12-12 всегда0. 8 http://habshuncoy.webcindario.com/bonoju/planirovanie-10-klass-russkiy-yazik-balandina-planirovanie.php2019-12-13всегда0,5http: //habshuncoy.webcindario.com/bonoju/vse-domashnie-zadaniya-zadanium по-русскому-языку-3-класс-впканакина-школа-россии-2020-год.php2019-12-14 всегда0.5 http://habshuncoy.webcindario.com/bonoju/gdz-po-russkomu-yaziku-10-klass-grekov-2020.php2019-12-15всегда0.6http: //habshuncoy.webcindario.com/ bonoju / gdz-russkiy-yazik-5-klass-lvova-2020-onlayn.php2019-12-16 всегда0.8 http://habshuncoy.webcindario.com/bonoju/gdz-po-russkomu-dlya-klassa-ta-ladizhenskaya. php2019-12-17всегда0,6 http://habshuncoy.webcindario.com/bonoju/reshebnik-po-fizike-7-9-klass-orlov-tatur.php2019-12-18всегда0,5 http://habshuncoy.webcindario.com/ боною / гдз-русский-4-класс-1-часть-зеленина-хохлова.php2019-12-19всегда0.8 http://habshuncoy.webcindario.com/bonoju/gdz-po-russkomu-yaziku-5-klass-vilenkin.php2019-12-20всегда0.5http: //habshuncoy.webcindario.com/bonoju/ диктанти-для-3-класс-по-русскому-языку-в-беларуси.php2019-12-22 всегда 0,7 http://habshuncoy.webcindario.com/bonoju/otveti-na-russkiy-yazik-2-klassa-zheltovskaya. php2019-12-23всегда0,7 http://habshuncoy.webcindario.com/caviho/always1 http://habshuncoy.webcindario.com/caviho/resheba-5-klass.php2019-10-25 всегда0,8 http://habshuncoy.webcindario.com / caviho / rabochaya-tetradi-po-geometrii-8-klass-atanasyan.php2019-10-27всегда0.7 http://habshuncoy.webcindario.com/caviho/bileti-i-otveti-po-biologii-9-klass. php2019-10-29всегда0.5 http://habshuncoy.webcindario.com/caviho/formula-po-fizike-9-klass-perishkin.php2019-10-31 всегда0.7 http://habshuncoy.webcindario.com/caviho/trenirovochnaya- работа-по-физике-8-октября-2020-года-9-класс-видеоразбор.php2019-11-02 всегда 0,7 http://habshuncoy.webcindario.com/caviho/rayonniy-monitoring-po-geometrii-9-klass. php2019-11-04всегда0.5 http://habshuncoy.webcindario.com/caviho/gdz-po-konturnim-kartam-po-geografii-7-klass-drofa.php2019-11-06 всегда0,8 http://habshuncoy.webcindario.com/caviho/atlas- po-geografii-5-klass-drofa.php2019-11-08 всегда0.5 http://habshuncoy.webcindario.com/caviho/rabochaya-tetral-4-klass-kuzovlev.php2019-11-10 всегда0.7http: // habshuncoy. webcindario.com/caviho/skachat-otveti-po-literaturnomu-chteniyu-3-klass-boykina.php2019-11-13 всегда0,5 http://habshuncoy.webcindario.com/caviho/testovie-zadaniya-po-obshestvoznaniyu-10 класс-1-полугодие.php2019-11-16всегда0,6 http://habshuncoy.webcindario.com/caviho/didakticheskie-materiali-po-himii-8-klass-gorshkova-s-temami.php2019-11-18 всегда0,6 http://habshuncoy.webcindario. com / caviho / gdz-po-leteraturnomu-chteniyu-4-klass-buneev.php2019-11-21always0.6http: //habshuncoy.webcindario.com/caviho/skachat-gdz-po-nemetskomu-za-10-11- klass.php2019-11-22всегда0.6 http://habshuncoy.webcindario.com/caviho/gdz-po-angliskomu-4-klass-znnikitenko.php2019-11-23всегда0.7http: //habshuncoy.webcindario/caviho.com/caviho тетрад-для-практических-работ-по-географии-9-класс-аг-стадник.php2019-11-24 всегда0,5 http://habshuncoy.webcindario.com/caviho/uchebniki-dlya-7-klassa-na-planshet.php2019-11-26 всегда0,7 http://habshuncoy.webcindario.com/caviho/dlya- 6-го-класс-сканворд-по-теме-культура-халифата-сканворд.php2019-11-28всегда0.5 http://habshuncoy.webcindario.com/caviho/reshenie-nomerov-po-algebre-8-klass-mordkovich. php2019-11-29 всегда0.5 http://habshuncoy.webcindario.com/caviho/spotlight-5-klass-workbook-audio.php2019-11-30 всегда0.6 http://habshuncoy.webcindario.com/caviho/spishi-ru- биология-9-класс.php2019-12-02 всегда0.6 http://habshuncoy.webcindario.com/caviho/gdz-po-rabochey-tetradi-pol-informatike-klass.php2019-12-03 всегда0.5 http://habshuncoy.webcindario.com/caviho/ решебник-по-литературному-чтению-2-класс-м-бойкина.php2019-12-04всегда0.7 http://habshuncoy.webcindario.com/caviho/radetskiy-didakticheskiy-material-gdz-10-klass-19-skachat.php 12-05всегда0.7 http://habshuncoy.webcindario.com/caviho/gdz-algebra-i-nachalo-matematicheskogo-analiza-10-klass-mordkovich-profilniy-uroven.php2019-12-06 всегда0.6 http://habshuncoy.webcindario.com/caviho/gdz-po-istorii-klass-aa-danilov.php2019-12-08 всегда0,6 http://habshuncoy.webcindario.com/caviho/students-book-9-klass- 16-izdanie.php2019-12-09 всегда0,6 http://habshuncoy.webcindario.com/caviho/rabochaya-tetrad-kaufman-7-klass.php2019-12-10 всегда0,6 http://habshuncoy.webcindario.com/caviho/ gdz-gotovie-domashnie-zadaniya-po-geometrii-za-klass.php2019-12-12 всегда 0.8 http://habshuncoy.webcindario.com/caviho/7-klass-gdz-po-algebre-mordkovich-i-dr. php2019-12-13всегда0.5 http://habshuncoy.webcindario.com/caviho/knigi-10-klass-skachat-na-planshet.php2019-12-14всегда0,8 http://habshuncoy.webcindario.com/caviho/plani-konspekti-urokov-po- obzh-9-klass.php2019-12-16 всегда0.8 http://habshuncoy.webcindario.com/caviho/skachat-testi-po-vseobshey-istorii-7-klass.php2019-12-16 всегда0.7 http://habshuncoy. webcindario.com/caviho/tetrad-dlya-laboratornih-rabot-po-biologii-7-klass-otveti-poperenko.php2019-12-17 всегда0,7 http://habshuncoy.webcindario.com/caviho/istoriya-5-klassa- все-ответы-в-тетради.php2019-12-18всегда0.6 http://habshuncoy.webcindario.com/caviho/gdz-po-gia-9-klass-pod-redaktsiey-fflisenko-syukulabuhova-2020g.php2019-12-19 всегда0.7http: // habshuncoy. webcindario.com/caviho/reshebnik-po-himii-10-klass-profilniy-uroven.php2019-12-20всегда0.8 http://habshuncoy.webcindario.com/caviho/gdz-po-angliyskumu-8-klass-pismo- drugu.php2019-12-22всегда0.6http: //habshuncoy.webcindario.com/caviho/reshebnik-umniki-i-umnitsi-3-klass.php2019-12-23всегда0.8http: //habshuncoy.webcindario.com/hytepur/ always1http: // habshuncoy.webcindario.com/hyhytepur/gdz-po-pechatnoy-tetradi-biologiya-7.php2019-10-25всегда0.7 http://habshuncoy.webcindario.com/hyhytepur/bzhd-uchebnik.php2019-10-27 всегда0.6http:// habshuncoy.webcindario.com/hyhytepur/skachat-uchebnik-8-klliteraturazvinyatskovskiy-filenko.php2019-10-29 всегда 0,6http://habshuncoy.webcindario.com/hyhytepur/aprimkevich-zbornik-gizadach php2019-10-31 всегда0,6 http://habshuncoy.webcindario.com/hyhytepur/biologiya-10-11-gdz.php2019-11-02 всегда0,8 http://habshuncoy.webcindario.com/hyhytepur/test-po-biologii-7-tip-chervi-proverka-svoih-znaniy.php2019-11-04 всегда0.6 http://habshuncoy.webcindario.com/hyhytepur/bendrikov-fizika-gdz-reshebnik- skachat.php2019-11-06 всегда0,7 http://habshuncoy.webcindario.com/hyhytepur/gdz-po-uchebniku-lg-peterson-gv-dorofeev.php2019-11-08 всегда0,8 http://habshuncoy.webcindario.com/ hyhytepur / gdz-po-algebre-alimov.php2019-11-10 всегда0.5 http://habshuncoy.webcindario.com/hyhytepur/urovni-srednevekovoy-kulturi-tablitsa.php2019-11-13 всегда0.5 http://habshuncoy.webcindario.com/hyhytepur/pnsh3klass-kontrolnaya-rabota-za-2-yu-chetvert.php2019-11-16 всегда0,5 http://habshuncoy.webcindario.com/hyhytepur/uchebniki-po-t pravu-skachat-2020-god-format-doc.php2019-11-19 всегда0.6 http://habshuncoy.webcindario.com/jywekux/always1http: //habshuncoy.webcindario.com/jywekux/nem-yaz-5-klas- rabochaya-tetrad-2020.php2019-10-25 всегда0,7 http://habshuncoy.webcindario.com/jywekux/gotov-zavdannya-z-konturno-karti-10-klas.php2019-10-27всегда0,5 http://habshuncoy.webcindario.com/jywekux/ya-hotel-chtob-v-serdtse-spominalo-sad-i-leto-gdz.php2019-10-29 всегда0,6 http://habshuncoy.webcindario.com/jywekux/obshaya-elektrotehnika-s- osnovami-elektroniki-otveti.php2019-10-31 всегда0.6 http://habshuncoy.webcindario.com/jywekux/gdz-evgladchenko-gakasyan-mapoznyak-diholin.php2019-11-02 всегда0.8http: //habshuncoy. jywekux / gdz-6-klas-ros-mova-balandina-degtereva-lebedenko.php2019-11-04 всегда0.8 http://habshuncoy.webcindario.com/jywekux/dudnitsi-yu-p-geometriya-gdz-kla.php2019-11-06 всегда0,6 http://habshuncoy.webcindario.com/jywekux/gotovie-domashnie-zadaniya-k-uchebniku-istorii.php2019-11-09 всегда0,5 http://habshuncoy.webcindario.com/jyve-kux/ot k-labratornoy-rabote-po-fizike-ilechenko-kulikovskiy-7klass.php2019-11-10всегда0.6 http://habshuncoy.webcindario.com/jywekux/gdz-6-klas-zagalna-geografya-oyaskuratovich.php2019-11-10 13всегда0,8 http://habshuncoy.webcindario.com/jywekux/latin-reshebnik-gorodkova.php2019-11-16 всегда0,6 http://habshuncoy.webcindario.com/jywekux/gdz-k-uchebniku-alimova-10-11- клас.php2019-11-19 всегда0,5 http://habshuncoy.webcindario.com/cikom/always1 http://habshuncoy.webcindario.com/cikom/gdz-7-klass-angliyskiy-nataliya-koltko-bogdan.php2019-10-26. 6 http://habshuncoy.webcindario.com/cikom/rabochaya-tetrad-po-geografii-8-klass-kim-otveti-slayd-shou-video.php2019-10-27всегда0,6http: //habshuncoy.webcindario.com/ cikom / informatika-7-klass-semakin-gdz.php2019-10-29always0.7 http://habshuncoy.webcindario.com/cikom/istoriya-za-7-klass-danilov-chitat.php2019-10-31always0.7http: // habshuncoy.webcindario.com/cikom/stranichki-lyuboznatelnih-moro-3-klass-igra-odinnadtsat-palochek.php2019-11-02 всегда0.7 http://habshuncoy.webcindario.com/cikom/pochva-vazhneyshaya-chast-ekosistemi-pre 3-klass.php2019-11-04 всегда0.8 http://habshuncoy.webcindario.com/cikom/po-uchebniku-fiziki-8-klassa-asperishkin-testi-ik-nemu-otveti.php2019-11-06 всегда0.8http: //habshuncoy.webcindario.com/cikom/gdz-klass-po-anglisskomu-yaziku.php2019-11-09 всегда0.5 http://habshuncoy.webcindario.com/cikom/konspekt-zadacha-nbistomina-2-klass.php2019-11-10 всегда 0,6 http://habshuncoy.webcindario.com/cikom/angliyskiy-5-klass-oksana-karpyuk-otveti-k-rabochey-tetradi.php2019-11-13 всегда 0,6 http://habshuncoy.webcindario. com / cikom / pomogite-perevesti-tekst-s-angliyskogo-na-russkiy-5-klass.php2019-11-16always0.6http: //habshuncoy.webcindario.com/cikom/istoriya-10-klass-saharov-1- chast-gdz.php2019-11-19 всегда0,8 http://habshuncoy.webcindario.com/cikom/otveti-na-krossvordi-vedyushkin-6-klass.php2019-11-21 всегда0,8 http://habshuncoy.webcindario.com/ циком / презентация-история-владимирского-край-класс.php2019-11-22всегда0,7 http://habshuncoy.webcindario.com/cikom/bilyanina-10-klass-gdz.php2019-11-23всегда0,8 http://habshuncoy.webcindario.com/cikom/gdz-po-geografii- k-paragrafam-10-klass-maksakovskiy.php2019-11-25 всегда0,5 http://habshuncoy.webcindario.com/cikom/gdz-po-geografii-klass-po-gerasimovoy-i-neklyukovoy.php2019-11-26 всегда0. 7 http://habshuncoy.webcindario.com/cikom/gdz-po-angliyskomu-11-klass-new-millenium-english.php2019-11-28 всегда 0,7 http://habshuncoy.webcindario.com/cikom/gdz-klass- 2020-года-випуска.php2019-11-29всегда0.6 http://habshuncoy.webcindario.com/cikom/gdz-po-obshestvoznaniyu-11-klass-ln-bogolyubova.php2019-12-01 всегда0.6 http://habshuncoy.webcindario.com/cikom краткий-консект-по-истории-россии-8-класс-данилов-параграфа-5.php2019-12-02всегда0.7 http://habshuncoy.webcindario.com/cikom/gdz-po-biologii-6-klass-paragraf- 18-тпо-пасечник-снисаренко-2020-onlain.php2019-12-03всегда0.7 http://habshuncoy.webcindario.com/cikom/olimpiada-po-obshestvu-za-7-klass-2020-god.php2019-12- 04всегда0,5 http://habshuncoy.webcindario.com/cikom/reshebnik-istoriya-drevnego-mira-rabochaya-tetrad-5-klass-1-chast-goder-zadanie.php2019-12-05 всегда0.7 http://habshuncoy.webcindario.com/cikom/reshenie- Упражнения-426-по-учебнику-6-класс-сильвова.php2019-12-06всегда0.6 http://habshuncoy.webcindario.com/cikom/gdz-po-angliyskomu-5-klass-kuzovlev-lapa-duvanova-kostina- kuznetsova.php2019-12-08 всегда0,6 http://habshuncoy.webcindario.com/cikom/gdz-po-angliyskomu-8-klass-mz-biboletova.php2019-12-09 всегда0,7 http://habshuncoy.webcindario.com / cikom / 7-класс-новая-история-8-9-параграф-художники-i-ih-proizvedeniya-baranov.php2019-12-10всегда0.8http: //habshuncoy.webcindario.com/cikom/uroki-angliyskogo- 3-класс-задание-24-номер-6-video.php2019-12-12всегда0.6 http://habshuncoy.webcindario.com/cikom/kompleksnaya-kontrolnaya-rabota-i-polugodie-2-klass-fgos.php2019- 12-14 всегда0,5 http://habshuncoy.webcindario.com/cikom/raznourovnevie-zadaniya-po-teme-reshenie-lineynih-neravenstv-9-klass.php2019-12-15 всегда0,7 http://habshuncoy.webcindario.com/ cиком / решебник-по-англ-яз-5-класс-вербицкая.php2019-12-16 всегда 0,8 http://habshuncoy.webcindario.com/cikom/perevod-teksta-s-angliyskogo-na-russkiy-8-klass-kaufman.php2019-12-16 всегда 0,8 http://habshuncoy.webcindario. com / cikom / konspekt-uroka-vostok-v-nachale-20-veka-9-klass-noveyshaya-istoriya.php2019-12-17 всегда0.5 http://habshuncoy.webcindario.com/cikom/rabota-so-slovaryami- vo-2-klasse-pnsh.php2019-12-18 всегда0,8 http://habshuncoy.webcindario.com/cikom/gdz-po-uchebniku-geografii-klass.php2019-12-19 всегда0,6 http://habshuncoy.webcindario. ком / циком / плани-по-биологии-7-класс-латюшин.php2019-12-20всегда0,8 http://habshuncoy.webcindario.com/cikom/galitskiy-reshebnik-8-klass-21.php2019-12-22всегда0,8 http://habshuncoy.webcindario.com/cikom/skachat-gdz- по-истории-9-класс-загладин-всеобщая-история.php2019-12-23always0.5 http://habshuncoy.webcindario.com/cikom/perevod-teksta-po-angliyskomu-5-klass.php2019-12-24always0. 5 http://habshuncoy.webcindario.com/vuzoqozaw/always1http: //habshuncoy.webcindario.com/vuzoqozaw/gdz-algebra-klass-yun-makarichev-ng-mindyuk-ki-neshkov.php2019-10-26.5 http://habshuncoy.webcindario.com/vuzoqozaw/gdz-po-fizike-k-uchebniku-gladkova-i-tsodikova.php2019-10-27всегда0,8http: //habshuncoy.webcindario.com/vuzoqozaw-po/reshebniku prirodoznavstvo-5-rjnbr.php2019-10-29 всегда0.5 http://habshuncoy.webcindario.com/vuzoqozaw/gdz-po-algebre-8-klass-makarichev-pod-redaktsiey-telyakovskogo-17e-izdanie. 31всегда0,6 http://habshuncoy.webcindario.com/vuzoqozaw/gdz-sbornik-testov-ege-po-algebre.php2019-11-02 всегда0,7 http://habshuncoy.webcindario.com/vuzoqozaw/gdz-po-algedre классов-макаричев-14-издание.php2019-11-04всегда0.6 http://habshuncoy.webcindario.com/vuzoqozaw/reshebnik-po-algebre-9-klass-makarichev-2020-smotret.php2019-11-06 всегда0.6 http://habshuncoy.webcindario.com/ vuzoqozaw / search-gdz-po-program-razumovskoy.php2019-11-09 всегда0.5 http://habshuncoy.webcindario.com/vuzoqozaw/otveti-na-testi-po-fizike-9klass-gromtseva.php2019-11-10 всегда0. 5 http://habshuncoy.webcindario.com/vuzoqozaw/konspekt-uroka-po-sbo-na-temu-uhod-za-golovnimi-uborami.php2019-11-14 всегда0,6 http://habshuncoy.webcindario.com / vuzoqozaw / skachat-bez-registratsii-po-fgos-dlya-zabotlivih-roditeley-all-domashnie-zadaniya-2klassreshebnik-iv.php2019-11-16always0.7Продольный анализ SARS-CoV-2 на единичные клетки в эпителии дыхательных путей человека
Резюме
SARS-CoV-2, возбудитель COVID-19, трагически обременил людей и учреждения по всему миру. В настоящее время нет одобренных лекарств или вакцин для лечения или профилактики COVID-19. Углубленное понимание инфекции и патогенеза SARS-CoV-2 имеет решающее значение для разработки терапевтических средств.Чтобы получить представление о вирусной репликации, клеточном тропизме и взаимодействии вируса-хозяина SARS-CoV-2, мы выполнили секвенирование одноклеточной РНК экспериментально инфицированных эпителиальных клеток бронхов человека (HBEC) в интерфейсных культурах воздух-жидкость с течением времени. Это позволило выявить новые полиаденилированные вирусные транскрипты и выделить ресничные клетки как главную мишень инфекции, что мы подтвердили с помощью электронной микроскопии. В ходе инфекции клеточный тропизм SARS-CoV-2 распространяется на другие типы эпителиальных клеток, включая базальные и булавовые клетки.Инфекция индуцирует присущую клетке экспрессию IFN типа I и типа III и IL6, но не IL1. Это приводит к экспрессии генов, стимулированных интерфероном, как в инфицированных, так и в клетках-свидетелях. Мы наблюдаем аналогичные изменения экспрессии генов у пациента с COVID-19 ex vivo . Кроме того, мы разработали новый вычислительный метод, названный условным встраиванием плотности (CONDENSE), чтобы охарактеризовать и сравнить динамику темпоральных генов в ответ на инфекцию, который выявил гены, связанные с эндотелином, ангиогенезом, интерфероном и сигнальными путями, вызывающими воспаление.В этом исследовании мы провели углубленный анализ инфекции SARS-CoV-2 в HBEC и у пациента с COVID-19 и выявили гены, типы клеток и изменения состояния клеток, связанные с инфекцией.
1. Введение
В декабре 2019 года в Ухане, Китай, была обнаружена новая вирусная пневмония, теперь известная как коронавирусная болезнь 2019 (COVID-19) [ 1 ]. Тяжелый острый респираторный синдром (SARS) — коронавирус (CoV) -2, возбудитель COVID-19, стал причиной ошеломляющего и растущего числа инфекций и смертей во всем мире.В настоящее время нет одобренных лекарств или вакцин для лечения или профилактики COVID-19. Клинические проявления сильно варьируют от бессимптомной инфекции до острого респираторного дистресс-синдрома и смерти [ 2 ]. Углубленное понимание вирусного патогенеза на клеточном и молекулярном уровне имеет решающее значение для разработки прогностических инструментов и новых методов лечения.
CoV — это вирусы с оболочкой с положительно-смысловыми одноцепочечными геномами РНК размером от 26 до 30 т.п.н. [ 3 ].Ранее было идентифицировано шесть CoV человека: HCoV-NL63 и HCoV-229E, которые принадлежат к роду Alphacoronavirus; и HCoV-OC43, HCoV-HKU1, SARS-CoV и CoV с ближневосточным респираторным синдромом (MERS-CoV), которые принадлежат к роду Betacoronavirus [ 4 ]. За последние два десятилетия CoV стали серьезной проблемой общественного здравоохранения из-за потенциальной зоонозной передачи, о чем свидетельствует появление SARS-CoV в 2002 году, от которого заразилось 8000 человек во всем мире со смертностью 10-15%, и MERS-CoV в 2012 и 2019 годах, заразивший 2500 человек со смертностью 35%, а теперь и SARS-CoV-2 [ 5 ].Тканевый и клеточный тропизм являются ключевыми детерминантами вирусного патогенеза. Проникновение SARS-CoV-2 в клетки зависит от связывания белка вирусного шипа (S) с его родственным рецептором ангиотензин-превращающего фермента II (ACE2) на поверхности клетки [ 2 ]. ACE2 также является рецептором для SARS-CoV и HCoV-NL63, но эти вирусы вызывают совершенно разные заболеваемость и смертность, что указывает на неизвестные детерминанты патогенеза коронавируса [ 6 , 7 ]. Кроме того, протеолитическое праймирование белка S протеазами хозяина также имеет решающее значение для проникновения вируса [ 8 ].Трансмембранная клеточная сериновая протеаза типа II (TMPRSS2) используется SARS-CoV-2 для праймирования S-белка [ 9 , 8 , 10 , 11 ]. Это также используется SARS-CoV вместе с эндосомальными цистеиновыми протеазами катепсином B и L [ 12 , 13 ]. Предполагается, что другая протеаза хозяина, фурин, опосредует патогенез SARS-CoV-2; однако точная роль протеаз хозяина в проникновении SARS-CoV-2 еще предстоит определить [ 14 , 11 ].
SARS-CoV и MERS-CoV вызвали фатальную пневмонию, связанную с быстрой репликацией вируса, повышением провоспалительных цитокинов и инфильтрацией иммунных клеток [ 15 ]. Эти характеристики аналогичным образом наблюдаются при инфекции SARS-CoV-2. У пациентов с COVID-19 повышены уровни провоспалительных эффекторных цитокинов, таких как TNF , α , IL1B и IL6, а также хемокинов, таких как CCL2 и CXCL10, особенно у тех, кто находится в критическом состоянии [ 16 , 17 , 18 , 19 ].Эти исследования показывают, что за патогенез COVID-19 может быть ответственен чрезмерно обильный иммунный ответ, характеризующийся цитокиновым штормом, а не прямым повреждением, вызванным вирусом. Типы клеток и механизмы, лежащие в основе этого иммунного ответа, для SARS-CoV-2 неясны. Кроме того, было замечено, что возраст является сильным фактором риска более тяжелого заболевания (CDC MMWR). В Соединенных Штатах с 12 февраля по 16 марта 2020 г. уровень летальности для пациентов в возрасте 85 лет и старше составил 10,4− −27,3% по сравнению с 0.1 — −0,2% для пациентов 20 — −44 лет (CDC MMWR). Причина повышенного риска остается неизвестной.
Наши знания биологии и патогенеза SARS-CoV-2 неполны. Чтобы устранить этот пробел, мы выполнили одноклеточное (sc) секвенирование РНК (RNA-seq) на органотипических эпителиальных клетках бронхов человека (HBEC), инфицированных SARS-CoV-2. Эта система культивирования поддерживает дифференцировку эпителиальных клеток и имитирует ключевые аспекты эпителия слизистой оболочки. Используя scRNA-seq и электронную микроскопию, мы обнаружили, что реснитчатые клетки являются основной мишенью для инфекции SARS-CoV-2.В ходе инфекции клеточный тропизм SARS-CoV-2 распространился на другие эпителиальные клетки, включая базальные и булавовые клетки. Кроме того, инфекция SARS-CoV-2 вызвала внутреннюю экспрессию интерферонов типа I и типа III и IL6, но не IL1. Экспрессия гена, стимулированного интерфероном (ISG), наблюдалась как в инфицированных, так и в популяциях клеток-свидетелей. Мы также обнаружили, что гены, по-разному экспрессируемые между инфицированными и ресничными клетками-свидетелями, были обогащены возрастными генами, что является критическим фактором риска COVID-19.Здесь мы предоставляем подробный анализ инфекции SARS-CoV-2 в HBECs и в клетках у педиатрического пациента с COVID-19 во время пика и разрешения болезни. Мы выявили новые транскрипты SARS-CoV-2, клеточный тропизм, экспрессию генов-хозяев и состояние клеток, связанных с инфекцией.
2. Результаты
2.1. Динамика вирусной инфекции
Чтобы охарактеризовать взаимодействия SARS-CoV-2 с дыхательными путями человека, мы выполнили секвенирование одноклеточной РНК эпителия дыхательных путей, инфицированного SARS-CoV-2. Мы культивировали первичные HBEC на границе раздела воздух-жидкость (ALI) в течение 28 дней, а затем провоцировали апикальную поверхность эпителия 10 4 бляшкообразующих единиц (PFU) SARS-CoV-2 ().Экспоненциальная репликация вируса в ходе эксперимента была продемонстрирована с помощью qRT-PCR клеточного лизата для гена нуклеокапсида (N) SARS-CoV-2 (). Через 1, 2 и 3 дня после заражения (dpi) была получена суспензия отдельных клеток, и 3 ‘секвенирование одноклеточной РНК было выполнено на 77 143 клетках в четырех образцах клеток на образец со средним значением 31 383 прочтений на ячейку (, S1A). Чтобы определить клетки, инфицированные SARS-CoV-2, мы сопоставили считывания с вирусным эталонным геномом и количественно определили количество вирусных транскриптов для каждой клетки ().Мы определили продуктивно инфицированные клетки как клетки, содержащие не менее десяти вирусных транскриптов на клетку, что контролирует фон из-за несогласованных считываний в имитирующем образце. В соответствии с репликацией вирусного генома () мы наблюдали зависящее от времени увеличение численности инфицированных клеток с 1 до 3 точек на дюйм (, S1B).
scRNA-seq выявляет инфицирование HBEC SARS-CoV-2. А . Схема эксперимента. Эпителиальные клетки бронхов человека (HBEC) культивировали и инфицировали или нет (имитация) SARS-CoV-2.Инфицированные культуры собирали для scRNA-seq через 1, 2 и 3 дня после заражения (dpi). В . RT-qPCR в культуре HBEC для обнаружения вирусных транскриптов через 1 час после инфицирования (hpi), через 1, 2 и 3 дня после заражения (dpi) (копии / лунку). С . UMAP-визуализация количества генов scRNA-seq после пакетной коррекции. Каждая точка представляет собой ячейку, окрашенную по образцу. Д . Нормализованное количество вирусов в каждом состоянии. Для каждой клетки подсчет количества вирусов определяли путем сопоставления считываний с одним эталоном для всего генома. E . Процент клеток, инфицированных SARS-CoV-2, на основе порогового значения количества вирусных генов (см. Материалы и методы) F . Нормализованная тепловая карта количества вирусных открытых рамок считывания (ORF) в каждом состоянии. Считывания были сопоставлены с каждыми 10 ORF SARS-CoV-2. Г . График охвата вирусных считываний, выровненных с геномом SARS-CoV-2. Глубину секвенирования рассчитывали для каждой позиции в геноме для каждого условия. По мере развития инфекции охват генома становится более рассредоточенным. Н .Визуализация UMAP инфицированных клеток и клеток-свидетелей в каждом состоянии после пакетной коррекции. Клетки-свидетели определяются как клетки, которые остались неинфицированными в образцах HBEC, зараженных SARS-CoV-2.
Затем мы охарактеризовали транскриптом SARS-CoV-2 на одноклеточном уровне. В дополнение к считываниям, которые, как ожидается, будут выровнены непосредственно перед каноническим хвостом поли-A SARS-CoV-2, наши результаты показывают выравнивание дополнительных считываний в другом месте генома, что предполагает существование неканонических полиаденилированных субгеномных РНК (sgRNA) ( ,).Распределение полиаденилированных вирусных транскриптов сдвигается с 3 ’на 5’ во время инфекции (,, S1C). Картирование продуктивных инфицированных клеток выявляет множественные кластеры инфицированных клеток, которые со временем увеличиваются и не присутствуют в фиктивной выборке (). Используя RT-PCR, мы успешно проверили два уникальных пика, один, который отображен в середине области 1ab открытой рамки считывания (ORF), и второй пик, который отображен рядом с границей ORF6 (рис. S1D, верхняя панель). Наши результаты подтверждают, что продукты ОТ-ПЦР, соответствующие каждому из двух пиков, появляются после 2 точек на дюйм (фиг. S1D, нижняя панель, красные стрелки).Важно отметить, что отсутствие этих полос ОТ-ПЦР в ложных образцах и образцах с разрешением 1 dpi предполагает, что они не являются результатом неспецифического олиго-d (T) примирования клеточных или вирусных РНК. Мы включили два положительных контроля, усиливающих продукты ОТ-ПЦР увеличивающейся длины из канонического хвоста поли-A SARS-CoV-2 (рис. S1D, нижняя панель, зеленые стрелки). Эти полосы RT-PCR появляются уже с 1 dpi, специфичны для инфицированных клеток и имеют ожидаемую длину. Эти положительные контроли подтверждают, что мы способны улавливать известные полиаденилированные вирусные транскрипты с помощью этой стратегии прайминга ОТ-ПЦР.
2.2. Идентификация клеточного тропизма SARS-CoV-2
Дыхательные пути человека состоят из различных типов эпителиальных клеток с критическими функциями в газообмене, структуре и иммунитете. Мы стремились определить клеточный тропизм SARS-CoV-2 в бронхиальном эпителии, поскольку дыхательные пути являются важной мишенью вирусного патогенеза. Мы идентифицировали восемь основных кластеров, включающих канонические типы эпителиальных клеток: ресничные клетки, базальные клетки, клубные клетки, бокаловидные клетки, нейроэндокринные клетки, ионоциты и клетки пучка (S2A).Мы также наблюдали промежуточную популяцию клеток между базальными клетками и клубными клетками (BC / club), вероятно, представляющими базальные стволовые клетки, дифференцирующиеся в клубные клетки. Анализ дифференциально экспрессируемых генов показывает, что эти кластеры клеток экспрессируют классические типоспецифические маркеры эпителиальных клеток (). Картирование инфицированных вирусом клеток в пределах определенных типов эпителиальных клеток показывает, что ресничные, базальные, булавовые и BC / Club-клетки восприимчивы к инфекции SARS-CoV-2, тогда как бокаловидные, нейроэндокринные, пучковые клетки и ионоциты относительно устойчивы к инфекции (,).При 1 dpi ресничные клетки составляют 83% инфицированных клеток и продолжают составлять большинство инфицированных клеток на протяжении всей инфекции (,). Однако во время продуктивной инфекции количество инфицированных базальных, булавовых и BC / клубных клеток также увеличивается, что позволяет предположить, что эти клетки являются основными вторичными мишенями (,). Распределение полиаденилированных вирусных транскриптов по длине генома сходно для разных типов инфицированных клеток (рис. S2C).
Тропизм клеток SARS-CoV-2. А . Визуализация UMAP кластеров ячеек с ручными аннотациями.Клетки сначала были кластеризованы с помощью алгоритма Лувена, а затем аннотированы в соответствии с панелью маркерных генов. В . Скрипичный график генов-маркеров аннотаций и предполагаемых релевантных генов SARS-CoV-2, основанный на недавней литературе. С . Визуализация аппроксимации и проекции унифицированного коллектора (UMAP) нормализованных количеств считываний SARS-CoV-2. Здесь чтения определялись как в. Д . Доля инфицированных клеток по условиям и типам клеток. E . Гистограмма количества инфицированных клеток на тип клеток в разных условиях. Ф . Оценка инфицирования, полученная из Manifold Enhancement of Latent Dimensions (MELD), показывающая степень инфицирования на клетку, стратифицированную по времени клетки. Г . Изображение с помощью просвечивающей электронной микроскопии имитаций (слева) и SARS-CoV-2 эпителиальных клеток бронхов человека (HBEC) выявляет инфицированные реснитчатые клетки через 2 дня после инфицирования (dpi) (справа). Масштаб (внизу) соответствует 500 нм. Красными стрелками обозначены вирусные частицы, а черными — реснички.
Для независимой проверки тропизма клеток SARS-CoV-2, HBEC, культивированные в тех же условиях, что и scRNA-seq, оценивали с помощью просвечивающей электронной микроскопии.При 2 dpi мы наблюдали многочисленные вирусные частицы размером около 80 нм в реснитчатых клетках (). Это согласуется с известным распределением коронавирусов по размерам [ 20 ]. Эти частицы не наблюдались в имитационном контрольном образце (). В совокупности это подтверждает, что реснитчатые клетки являются основной мишенью инфекции SARS-CoV-2 в бронхах человека.
2.3. Детерминанты клеточного тропизма
Затем мы попытались определить транскрипционные корреляты хозяина тропизма клеток SARS-CoV-2.Поскольку проникновение вируса является основной детерминантой клеточного тропизма, мы сначала исследовали, предсказывает ли экспрессия рецептора SARS-CoV-2 ACE2 инфекцию. Мы наблюдали экспрессию ACE2 на низких уровнях в реснитчатых, базальных, булавовых и BC / булавных клетках в имитационном состоянии (). Неожиданно, экспрессия ACE2 плохо коррелировала с инфекцией SARS-CoV-2 на клеточной основе (r Спирмена между вирусным геномом и ACE2 в реснитчатых клетках, -0,06, и между ACE2 и оценкой инфекции в мерцательных клетках, 0,09). Однако экспрессия ACE2 была увеличена в четырех популяциях восприимчивых клеток: реснитчатых, базальных, булавовых и BC / клубных по сравнению с типами нечувствительных клеток: нейроэндокринными, ионоцитами, бокаловидными и пучковыми клетками (рис. S3).Экспрессия ACE и CLTRN, которые являются структурными гомологами ACE2 и аминопептидаз ANPEP и DPP4 (рецептор MERS-CoV), также слабо коррелировала с восприимчивостью к SARS-CoV-2 (-).
Экспрессия известных детерминант входа в типы бронхиальных эпителиальных клеток. А . Визуализации UMAP, окрашенные экспрессией четырех рецепторов и протеаз, экспрессируемых в эпителиальных клетках бронхов человека (HBEC): ACE2, TMPRSS2, TMPRSS4 и CTSL. B — E . Тепловые карты рецепторов и протеаз у реснитчатых ( B .), базальные ( C .), булавые ( D .) и BC / Club клетки ( E .).
ACE2 недавно продемонстрировал, что это ген, стимулированный интерфероном (ISG) [ 21 ]. Здесь мы наблюдаем умеренное увеличение экспрессии ACE2 как в инфицированных клетках, так и в клетках-свидетелях по сравнению с ложным образцом, что соответствует динамической регуляции экспрессии ACE2 с помощью врожденного иммунного ответа хозяина на SARS-CoV-2 (-). Чтобы проверить, объясняет ли экспрессия других потенциально провирусных генов тропизм клеток SARS-CoV-2, мы оценили экспрессию протеаз, которые могут усиливать инфекцию SARS-CoV-2.Трансмембранная сериновая протеаза TMPRSS2 и катепсин L участвуют в проникновении SARS-CoV-2 [ 22 ]. Мы также исследовали родственную протеазу TMPRSS4, которая расщепляет гемаглютинин гриппа, аналогично TMPRSS2, а также может играть роль в проникновении SARS-CoV-2 [ 22 , 23 ]. TMPRSS2 и CTSL экспрессировались преимущественно в базальных, клубных и реснитчатых клетках, в то время как TMPRSS4 широко экспрессировалась во всех типах эпителиальных клеток (-). Специфическая роль протеаз в управлении тропизмом SARS-CoV-2 в эпителии дыхательных путей человека требует дальнейшего изучения.
2.4. Врожденный иммунный ответ на инфекцию SARS-CoV-2
SWe исследовал транскриптом хозяина, чтобы оценить иммунный ответ хозяина на инфекцию SARS-CoV-2 при одноклеточном разрешении в эпителии дыхательных путей человека. Мы наблюдали устойчивую индукцию интерферона типа I (IFNB1) и интерферонов типа III (IFNL1, IFNL2 и IFNL3) в реснитчатых, базальных, клубных и BC / клубных клетках, коэкспрессирующих транскрипты SARS-CoV-2 (2). Интересно, что кинетика индукции IFNB1 была замедленной по сравнению с интерфероном III типа.Напротив, индукция IFN в неинфицированных реснитчатых, базальных, булавовых и BC / клубных клетках была незначительной. Мы также не наблюдали индукцию IFN в нейроэндокринных клетках, ионоцитах, бокаловидных или пучковых клетках, соответствующих этим типам клеток, которые не являются основными клетками-мишенями SARS-CoV-2 (рис. S4). Это демонстрирует, что прямая инфекция SARS-CoV-2 данной клетки имеет решающее значение для индукции интерферона. Интерфероны типа I и III передают сигнал через IFNAR и IFNLR соответственно, что приводит к экспрессии сотен ISG. В соответствии с этим, мы наблюдали широкую индукцию ISG (IFI27, IFITM3, IFI6, MX1 и ISG15) как в инфицированных, так и в случайных клетках всех типов клеток (, S4), предполагая, что IFN из инфицированных клеток действует в транс- на обе инфицированные клетки. и незараженные случайные клетки.
Инфекция SARS-CoV-2 вызывает устойчивый врожденный иммунный ответ. A — D . Тепловые карты экспрессии ключевых генов врожденного иммунитета и воспаления в ресничных ( A .), Базальных ( B .), Клубных ( C .) И BC / клубных клетках ( D .). -регулируют интерфероны I и III типа, IL-6 и хемокины внутренним образом, в то время как гены, стимулированные интерфероном, индуцируются как в инфицированных, так и в клетках-свидетелях. Инфекция также стимулирует секрецию белка IL-6.Наблюдаются минимальные изменения в экспрессии и секреции белка IL-1A, IL-1B и IL1RN.
Противовирусный ответ хозяина также приводит к индукции хемокинов, что приводит к привлечению иммунных клеток, что является признаком тяжелой формы COVID-19 [ 24 ]. Здесь мы наблюдаем индукцию CXCL9, CXCL10 и CXCL11, которые распространяют сигналы через родственный рецептор CXCR3 для рекрутирования активированных Т-клеток и NK-клеток (). Эта индукция была очевидна в инфицированных, но не в случайных клетках (). Напротив, CCL2 и CXCL16, которые привлекают моноциты и Т-клетки, соответственно, не регулировались динамически в оцениваемых условиях (и S4).Мы также наблюдали существенную индукцию провоспалительного цитокина IL-6 в инфицированных ресничных, базальных, булавовых и BC / клубных клетках, но не в неинфицированных клетках-свидетелях этих же популяций. Интересно отметить, что экспрессия провоспалительного IL-1 была умеренно подавлена во всех типах клеток после инфицирования, тогда как экспрессия IL-10 и TNF α не регулировалась в значительной степени инфекцией в этой системе (2). Далее мы охарактеризовали уровни выбранных белков в базолатеральном супернатанте ложных и инфицированных HBEC и наблюдали индукцию IL6 и CXCL9, но не IL1A, IL1B и IL1RN, что согласуется с изменениями экспрессии генов ().
2.5. Дифференциально экспрессируемые гены в ответ на инфекцию SARS-CoV-2
Чтобы определить, как инфекция SARS-CoV-2 нарушает клеточный транскриптом, мы с помощью вычислений объединили три инфицированных образца и проанализировали 100 наиболее дифференциально экспрессируемых генов между инфицированными и неинфицированными клетками-свидетелями заданный тип ячеек в выборках с разрешением 1, 2 и 3 dpi (). Анализ онтологии генов PANTHER показал, что инфицированные реснитчатые клетки обладают повышенной экспрессией генов, участвующих в апоптозе (например,грамм. PMAIP1, SQSTM1, ATF3), инициацию трансляции и экспрессию вирусных генов (например, RPS12, RPL37A) и воспаление (например, NFKBIA и NFKBIZ) по сравнению с клетками-свидетелями (и списки дифференциально экспрессируемых генов в дополнительных файлах). Подобные гены обогащены в других популяциях инфицированных клеток (Рис. S5). Напротив, инфицированные ресничные клетки показали значительное подавление генов, включенных в биологические процессы, участвующие в функции ресничек (например, DYNLL1), передаче сигналов кальция (например, CALM1, CALM2) и гомеостазе железа (например,грамм. FTh2, FTL; , и S5).
Экспрессия дифференциально экспрессируемых генов. ( A ) Схема анализа дифференциальной экспрессии. Наблюдают две основные популяции клеток: клетки-свидетели, которые не были инфицированы вирусом через 3 дня после заражения (dpi), и инфицированные клетки, которые содержат активную репликацию вируса и транскрипцию при 3 dpi. ( B ) Графики вулкана, выделяющие наиболее дифференцированно экспрессируемые гены между инфицированными клетками и клетками-свидетелями в реснитчатых клетках с разрешением 3 точек на дюйм, измеренным с помощью расстояния земледельца (EMD).( C ) Тепловая карта наиболее дифференциально экспрессируемых генов между неинфицированными, инфицированными и случайными клетками в реснитчатых клетках во всех условиях. ( D ) Вулканический график дифференциально экспрессируемых генов в реснитчатых клетках (инфицированных против сторонних наблюдателей). Связанные с возрастом гены в легких человека имеют цветовую кодировку (синий, увеличение экспрессии с возрастом; оранжевый, снижение экспрессии с возрастом. ( E ) Диаграмма Венна, выделяющая пересечение между генами, связанными с возрастом легких, и SARS-CoV-2). регулируемые гены в ресничных клетках.Статистическая значимость перекрытия оценивалась гипергеометрическим тестом. ( F ) Характеристики возрастных генов, пораженных инфекцией SARS-CoV-2 в реснитчатых клетках. Статистическая значимость взаимодействия между направленностью регуляции SARS-CoV-2 (индуцированной или подавленной) и направленностью возрастной ассоциации (увеличение или уменьшение с возрастом) оценивалась с помощью точного двустороннего критерия Фишера. ( G ) Онтология гена DAVID и анализ путей генов, репрессированных инфекцией SARS-CoV-2 в реснитчатых клетках, экспрессия которых также снижается с возрастом.
Учитывая, что возраст является основным фактором риска серьезности COVID-19, мы исследовали, может ли экспрессия этих дифференциально экспрессируемых генов естественным образом меняться с возрастом [ 25 ]. Мы проанализировали дифференциально экспрессируемые гены между инфицированными и ресничными клетками-свидетелями для ранее определенных возрастных генов () [ 26 ]. Возрастные гены были значительно обогащены среди по-разному экспрессируемых генов (). Из этих генов на пересечении гены, репрессированные инфекцией SARS-CoV-2, имели тенденцию к снижению экспрессии с возрастом, и наоборот ().Анализ генов, которые подавляются инфекцией SARS-CoV-2 и экспрессия которых снижается с возрастом, продемонстрировала обогащение для нескольких онтологий и путей генов, в первую очередь путей, связанных с митохондриями (). В совокупности это говорит о том, что инфекция SARS-CoV-2 перепрограммирует клеточный транскриптом, что приводит к развитию вирусной инфекции и потенциально приводит к клеточной дисфункции и апоптозу, что способствует патогенезу COVID-19.
2.6. Динамика генов в инфицированных клетках и клетках-свидетелях с использованием CONDENSE
Чтобы исследовать динамику транскрипции в ответ на инфекцию SARS-CoV-2, мы разработали новый вычислительный метод для характеристики генов по их временной динамике (методы), названный встраиванием условной плотности (или CONDENSE) .Вкратце, CONDENSE использует скрытое (или псевдо) временное измерение и условную оценку плотности ядра для количественной оценки динамики по оси времени для каждого гена. В текущем приложении это делается отдельно для инфицированных и сторонних клеток. Затем CONDENSE вычисляет расстояния между оценками плотности, обеспечивая метрическое пространство, которое можно использовать для обучения многообразию. Результатом является встраивание генов в соответствии с их динамикой, которое впоследствии может быть использовано для последующего анализа, такого как визуализация, кластеризация и архетипический анализ.Здесь мы используем архетипический анализ для характеристики многообразия динамики генов и наблюдаем, что между инфицированными клетками и клетками-свидетелями, гены могут демонстрировать сходную динамику во времени (например, Архетипы 7), в то время как другие демонстрируют дифференциальную динамику (например, Архетипы 1, 3, 7, 18) (, S6). Отдельные гены могут быть представлены как комбинация этих архетипических динамик и сгруппированы по архетипам (,). В архетипах с дифференциальной динамикой во времени мы обнаруживаем, что транскрипты, являющиеся прототипами этих архетипов, участвуют в пути EGFR (EGFR, RRAS), ангиогенезе (RHOB, NOTCh2), передаче сигналов эндотелина (END2, PIK3R1), воспалении (RRAS, SHC1). ), передачу сигналов интерлейкина (MKNK2, IRS1) и передачу сигналов интерферона γ (ITGB1).Мы также наблюдаем различия между инфицированными клетками и клетками-свидетелями: EGFR, NOTCh2, RRAS, IRS1, PIK3R1, ITGB1 и SHC1 демонстрируют повышенную экспрессию в более поздние сроки в клетках-свидетелях, в то время как RHOB и NFKB2 демонстрируют повышенную экспрессию в более поздние сроки в инфицированных клетках. В обеих популяциях экспрессия MKNK2 и EDN2 со временем снижается. В генах, представляющих особый интерес для COVID-19, TMPRSS2, IL1RN и IL1A имеют временную динамику, наиболее типичную для архетипа 1 (снижение экспрессии с течением времени как в инфицированных, так и в клетках-свидетелях), в то время как IFITM3 является наиболее прототипом для архетипа 3 (увеличение экспрессии в инфицированных клетках). через некоторое время) ().Таким образом, CONDENSE, используемый для оценки динамики генов через 3 дня после инфицирования в HBEC, подчеркивает дивергенцию инфицированных генов и генов-свидетелей в ключевых сигнальных путях, связанных с иммунным ответом и регуляцией.
Транскриптомная динамика во времени в инфицированных клетках и клетках-свидетелях. ( A ) Обзор метода встраивания условной плотности (CONDENSE) для характеристики динамики транскрипции во времени ( B ) Кластеризация генов по их динамической, совместной плотности, стратифицированная инфицированными и случайными клетками, с использованием архетипического анализа.( C ) Изображения показывают условные оценки плотности архетипов, идентифицированных в генах, встроенных в пространство UMAP. Диаграммы разброса показывают соответствие LOESS для экспрессии генов по сравнению с предполагаемым временем для различных генов в архетипических кластерах, которые демонстрируют дифференциальную динамику во времени. ( D ) Загрузка представляющих интерес вирусных и иммунных генов в архетипы. ( E ) Скорость РНК представляющих интерес генов, окрашенная в зависимости от статуса инфекции для отдельных клеток. ( F ) Скорости РНК соответствуют отдельно для инфицированных клеток и клеток-свидетелей, выделяя интересующие гены и представления латентного времени, полученные из скорости РНК.
2.7. Динамика заражения через скорость РНК
Затем, чтобы исследовать динамику транскрипции в масштабе времени в часах, мы проанализировали скорость РНК отдельных клеток, подгоняя динамическую модель состояния транскрипции на основе количества сплайсированных и несплайсированных транскриптов () [ 27 ]. Путем встраивания скоростей РНК в UMAP для всех образцов мы идентифицировали две отдельные популяции инфицированных клеток — одну в высоком переходном состоянии, что соответствует высокоскоростным потокам оттока, а другую — в конечном состоянии.Ресничные клетки также существуют в двух скоростных состояниях РНК: одно в конечной точке более транскрипционно отличается, чем инфицированные клетки, а другое — в устойчивом состоянии, более транскрипционно сходное с инфицированными клетками. IL6 в инфицированных клетках находится либо в индуцированном, либо в подавленном состоянии, в то время как несколько инфицированных клеток экспрессируют IL6 в устойчивом состоянии (). ACE2 в клетках-свидетелях, по-видимому, преимущественно находится в индуцированном состоянии, в то время как TMPRSS2 и TMPRSS4 имеют умеренные скорости РНК, при этом самые высокие транскрипционные изменения происходят в базальных или клубных клетках.Использование скорости РНК для вычисления латентного времени подтверждает, что инфицированные реснитчатые клетки достигают конечной точки, которая также, по-видимому, имитирует транскрипционный переход инфицированных базальных клеток (). Экспрессия IL1RN как в инфицированных, так и в базальных клетках-свидетелях увеличивается со временем и существует преимущественно в устойчивом состоянии, тогда как IFIT1, MX1 и IFI27 имеют высокие скорости в индуцированном состоянии транскрипции в инфицированных реснитчатых клетках, но в подавленном состоянии в промежуточных реснитчатых клетках. При подборе отдельных моделей для инфицированных клеток и клеток-свидетелей наиболее динамичные гены в инфицированных клетках участвуют в воспалении (C5AR1, NFKBIA), передаче сигналов интерлейкина (IRS1) и передаче сигналов EGFR (EGFR, EREG).Передача сигналов EGFR и NFKBIA индуцируются на ранних этапах и более высоко экспрессируются на ранних этапах латентного времени, тогда как C5AR1 и IRS1 более высоко экспрессируются в более поздние моменты времени в инфицированных клетках. Напротив, большинство динамических генов клеток-свидетелей вовлечены в передачу сигналов, стимулированную ангиотензином II (PRKCA), апоптоз (MAP3K5), передачу сигналов EGFR и воспаление (ITGB1). В клетках-свидетелях большинство этих генов сильно экспрессируются в более позднее время. В совокупности скорость РНК подтверждает, что некоторые из наиболее динамичных транскриптов участвуют в регуляции иммунной системы, и подчеркивает, что гены, идентифицированные как дифференциально экспрессируемые в данных по экспрессии генов, также связаны с большими скоростями РНК в инфицированных реснитчатых клетках.
2,8. Клеточный тропизм в клиническом эндотрахеальном аспирате COVID-19
Чтобы сравнить наши результаты с in vivo контекстом, мы выполнили scRNA-seq на эндотрахеальном аспирате пациента с COVID-19 во время интубации (пик заболевания) и после экстубации (при разрешении болезни), а также на контрольном образце (см. Методы). В этих образцах, помимо эпителиальных, были идентифицированы различные иммунные клетки (,). По сравнению с контрольным пациентом у пациента с COVID-19 было более высокое содержание нейтрофилов и моноцитов (рисунок S7A).Были идентифицированы клетки, экспрессирующие по крайней мере один вирусный транскрипт из эндотрахеального аспирата, взятого во время интубации, но не было идентифицировано инфицированных клеток из экстубированного образца, что соответствует экстубации после клинического выздоровления. В частности, были идентифицированы ресничные клетки и обнаружено, что они экспрессируют вирусные транскрипты в интубированном образце эндотрахеального аспирата). Однако клетки альвеолярного типа I были наиболее сильно инфицированы процентным соотношением идентифицированных клеток с вирусным транскриптом (). Эпителиальные клетки в интубированном образце показывают повышенную экспрессию регуляторных генов иммунной системы в соответствии с данными in vitro HBEC (,,,).Кроме того, TMPRSS2, TMPRSS4 и ACE2 экспрессируются выше в эпителиальных эндотрахеальных аспиратных клетках интубированного образца по сравнению с экстубированным образцом (рис. S7). Однако экспрессия IL1 и IL6 выше в экстубированном образце по сравнению с интубированным образцом. Повышенную экспрессию IL1 и IL6 в экстубированном образце можно объяснить развитием у пациента вторичной пневмонии. Однако диагностика вторичной пневмонии у интубированных и тяжелобольных педиатрических пациентов неточна.Тем не менее, интубированного пациента лечили антибиотиками, что может объяснить расхождение между HBEC и клиническими образцами.
Динамика транскрипции SARS-CoV-2 в дыхательных путях у детей с COVID-19. ( A ) Единичные клетки из эндотрахеальной аспирационной жидкости здорового контроля и педиатрического пациента с COVID-19 во время интубации и экстубации. ( B ) Типы клеток, идентифицированные в эндотрахеальных образцах. ( C ) Изобилие различных типов клеток в каждом клиническом образце.( D ) Процент клеток на тип клеток с хотя бы одним считыванием, сопоставленным с вирусным геномом. ( E ) Экспрессия представляющих интерес генов иммунной системы в эпителиальных клетках из ЖБАЛ у детей. ( F ) Экспрессия 25 лучших дифференциально экспрессируемых генов между интубированными и экстубированными образцами среди эпителиальных клеток из педиатрических образцов. ( G ) Дифференциальная экспрессия гена для распределительного расстояния по сравнению с скорректированным значением p для эпителиальных клеток в педиатрическом эндотрахеальном аспирате.
3. Обсуждение
Чтобы эффективно лечить COVID-19, мы должны сначала понять, как SARS-CoV-2 вызывает заболевание и почему клинические проявления варьируются от бессимптомной инфекции до смертельного заболевания. Здесь мы сообщаем о первом продольном одноклеточном транскриптомном анализе респираторного эпителия, инфицированного SARS-CoV-2, с использованием органоидной модели, которая воспроизводит ключевые свойства ориентации эпителия дыхательных путей. Генерируемые транскрипционные данные высокого качества, в среднем от 2400 до 3600 уникальных генов, обнаруживаемых для каждого состояния.Наши данные показывают несколько новых вирусных транскриптов, и наша методология позволяет дифференцировать инфицированные клетки от клеток-свидетелей. Кроме того, мы демонстрируем, что реснитчатые клетки являются основными клетками-мишенями инфекции SARS-CoV-2 в эпителии бронхов в начале инфекции и что клеточный тропизм со временем распространяется на базальные, булавы и BC / булавы клетки. Мы также обнаружили, что SARS-CoV-2 сильно индуцирует IFN в инфицированных клетках, что приводит к широкой экспрессии ISG как в инфицированных, так и в клетках-свидетелях. Мы также наблюдаем сильную индукцию провоспалительного цитокина IL-6 и хемокинов, которые, вероятно, способствуют воспалительной реакции in vivo .Кроме того, мы обнаружили, что гены, по-разному экспрессируемые между инфицированными и ресничными клетками-свидетелями, обогащены возрастными генами, что свидетельствует о потенциальном механизме увеличения тяжести заболевания у пожилых пациентов.
Одноклеточная транскриптомика позволила нам выяснить транскриптом SARS-CoV-2 при разрешении одной клетки во множестве первичных типов клеток с течением времени. Мы разработали новый метод дифференциации типов продуктивно инфицированных клеток по распределению субгеномных и геномных вирусных транскриптов.Мы также идентифицировали полиаденилированные вирусные транскрипты, удаленные от 3 ’конца вирусного генома, что было неожиданно с учетом нашего метода секвенирования. Наши валидационные эксперименты с ОТ-ПЦР подтверждают получение по крайней мере двух уникальных S-независимых транскриптов (регуляторная последовательность транскрипции (TR) S-независимых транскриптов с поли-A-хвостами, которые, по-видимому, не являются результатом неспецифического праймирования олиго-d (T). сообщенная скорость рекомбинации для коронавирусов высока [ 28 , 29 ], возможно, эти короткие чтения соответствуют неспецифическому скачку полимеразы.Однако недавние исследования выявили TRS-независимые химерные РНК, продуцируемые во время инфицирования SARS-CoV-2 клеток Vero, небольшая часть (1,5%) из которых слита в рамке [ 30 ]. Взятые вместе с нашими результатами, это может указывать на то, что неканонические субгеномные РНК с кодирующим потенциалом образуются во время инфекции SARS-CoV-2; однако это потребует дополнительной проверки. Недавно было показано, что неструктурный белок 8 (nsp8) HCoV-299E обладает матрично-независимой аденилтрансферазной активностью [ 31 ].Поскольку поли-A-хвосты играют важную роль в стабильности и потенциале трансляции канонических sgRNA SARS-CoV-2, интересно предположить, что коронавирусы могут полагаться на продукцию неканонических полиаденилированных sgRNA, которые служат приманками для клеточных деаденилазы. Это привело бы к сохранению поли-А-хвостов геномных и субгеномных РНК. В самом деле, продукция sgRNA во время флавивирусных инфекций важна для устойчивости к клеточным экзорибонуклеазам и уклонения от врожденного иммунитета [ 32 , 33 ].
Идентификация типов клеток, инфицированных SARS-CoV-2, определяет патогенез. Мы обнаружили, что SARS-CoV-2 в первую очередь поражает реснитчатые клетки, базальные, булавы и BC / булавы клетки. Это может привести к нарушению функции этих критических типов клеток. Ресничные клетки, которых много в респираторном эпителии, отталкивают слизь и связанные с ней инородные частицы и микробы от нижних дыхательных путей. Наше открытие, что реснитчатые клетки являются преобладающими клетками-мишенями инфекции SARS-CoV-2 в бронхиальном эпителии, имеет несколько важных последствий.Во-первых, дисфункция реснитчатых клеток в результате инфицирования SARS-CoV-2 может ухудшить мукоцилиарный клиренс и повысить вероятность вторичной инфекции. Во-вторых, астма, хроническая обструктивная болезнь легких и курение связаны как с дисфункцией ресничек, так и с повышенной тяжестью COVID-19. Изменяют ли эти основные состояния реснитчатые клетки и тем самым повышают их восприимчивость к инфекции, остается неясным, хотя SARS-CoV-2 был идентифицирован в реснитчатых клетках госпитализированных пациентов, что подчеркивает потенциальную связь между восприимчивостью реснитчатых клеток к инфекции SARS-CoV-2. и прогрессирование заболевания COVID-19 [ 34 ].В-третьих, изменения экспрессии генов, вызванные инфекцией SARS-CoV-2 в реснитчатых клетках, отражают естественные изменения экспрессии, которые происходят с возрастом, потенциально обеспечивая понимание того, почему у пожилых пациентов ухудшаются исходы болезни. Клеточный тропизм SARS-CoV-2 в носовом эпителии и нижних дыхательных путях остается важной областью будущих исследований, которые еще больше улучшат наше понимание патогенеза COVID-19.
Заболевание у пациентов с COVID-19 характеризуется задержкой после передачи с появлением симптомов на седьмой день и пиком тяжести заболевания через 14 дней после заражения [ 35 , 36 ].Это отличается от сезонных коронавирусов человека и предполагает важную роль иммунного ответа хозяина в прогрессировании COVID-19. Несколько недавних исследований показали, что индукция врожденного иммунитета во время инфекции SARS-CoV-2 зависит от кинетики вирусного роста и множественности инфекции. [ 37 , 38 ]. Здесь мы показываем, что врожденный ответ на SARS-CoV-2 является интактным и быстрым, что характеризуется индукцией интерферона, хемокина и IL-6. Интересно, что хотя мы не наблюдаем значительного истощения популяций чувствительных к вирусу клеток, мы обнаруживаем повышенную экспрессию генов, связанных с гибелью клеток, что предполагает, что антивирусный ответ хозяина является цитотоксическим и может вносить вклад в патогенез заболевания.В соответствии с этим, IL-6 является мощным провоспалительным цитокином, а уровни IL-6 в сыворотке позволяют прогнозировать дыхательную недостаточность [ 39 ]. Терапия, направленная на рецептор IL-6, в настоящее время проходит клинические испытания для лечения COVID-19. Эта работа поднимает ряд важных будущих направлений, включая то, взаимодействуют ли другие дыхательные пути и эндотелиальные ткани аналогичным образом с SARS-CoV-2 и как эти взаимодействия меняются in vitro .
4. Методы
4.1. Культура поверхности раздела воздух-жидкость HBEC
HBEC, от Lonza, культивировали в суспензии в среде PneumaCult-Ex Plus в соответствии с инструкциями производителя (StemCell Technologies, Кембридж, Массачусетс, США).Для создания культур поверхности раздела воздух-жидкость HBEC помещали на покрытые коллагеном вставки трансвелл с размером пор 0,4 микрон (Costar, Corning, Tewksbury, MA, USA) при 5 × 10 4 клеток / мл на фильтруют и помещают в 24-луночные культуральные планшеты. Клетки поддерживали в течение первых 3 дней в среде PneumaCult-Ex Plus, затем меняли на среду PneumaCult-ALI (StemCell Technologies), содержащую ингибитор ROCK Y-27632, в течение 4 дней. Свежая среда, 100 мкл в апикальной камере и 500 мкл в базальной камере, подавалась каждый день.На 7 день среду из апикальных камер удаляли, а в базальные камеры поддерживали 500 мкл среды PneumaCult-ALI. HBEC поддерживали на границе раздела воздух-жидкость в течение 28 дней, позволяя им дифференцироваться. Среду в базальной камере меняли каждые 2–3 дня (500 мкл ).
4.2. Вирусная инфекция
Изолят SARS-CoV-2 USA-WA1 / 2020 был получен из репозитория реагентов BEI. Все эксперименты по заражению проводились в учреждении с уровнем биобезопасности 3, лицензированным штатом Коннектикут и Йельским университетом.Непосредственно перед инфицированием апикальную сторону культуры HBEC ALI осторожно трижды промывали 200 мкл фосфатно-солевого буфера без двухвалентных катионов (PBS — / -). 10 4 бляшкообразующих единиц (PFU) SARS-CoV-2 в 100 мкл общего объема PBS добавляли в апикальный отсек. Клетки инкубировали при 37 C и 5% CO 2 в течение 1 часа. Несвязанный вирус удаляли, и клетки культивировали на границе раздела воздух-жидкость в течение до трех дней.Инфекции были перенесены на один день, и все четыре образца были обработаны одновременно для секвенирования одноклеточной РНК, как описано ниже.
4.3. Подготовка образцов для секвенирования одноклеточной РНК
Инокулированные HBEC промывали 1X PBS — / — и трипсинизировали ферментом TrypLE Express (ThermoFisher, Waltham, MA, USA) для получения суспензий одноклеточных. 100 мкл TrypLE добавляли в апикальную камеру, инкубировали в течение 10 минут при 37 ° C в инкубаторе CO 2 и осторожно пипетировали вверх и вниз для диссоциации клеток.Собранные клетки переносили в стерильную пробирку на 1,5 мл и нейтрализовали DMEM, содержащей 10 процентов FBS. Дополнительные 100 мкл TrypLE помещали в апикальную камеру, повторяя ту же процедуру, что и выше, в течение всего 30 минут для максимального сбора клеток. Клетки центрифугировали при 300 × g в течение 3 мин и ресуспендировали в 100 мкл DMEM с 10% FBS. Количество клеток и их жизнеспособность определяли с использованием исключения красителя трипанового синего на приборе Countess II (ThermoFisher Scientific).Целевой ввод клеток составлял 10 000 клеток на условие. Набор Chromium Next GEM (Gel Bead-In Emulsion) Single Cell 3 ’Gel beads v3.1 (10X Genomics, Pleasanton, CA, USA) использовали для создания GEM в соответствии с инструкциями производителя. Все образцы и реагенты были подготовлены, загружены в чип и запущены в Chromium Controller для создания GEM и штрих-кодирования. Созданные GEM использовали для синтеза кДНК и подготовки библиотеки с использованием набора Chromium Single Cell 3 ’Library Kit v3.1 (10X Genomics) в соответствии с инструкциями производителя.Сгенерированные библиотеки секвенировали в системе NovaSeq 6000 с использованием считываний пар оснований HiSeq 100 и двойного индексирования. Клетки секвенировали до средней глубины 31 383 считывания на ячейку. Геном человека, Ensembl GRCh48.98.gtf, и геном SARS-CoV-2, доступ к базе данных NCBI Genome {«type»: «entrez-nucleotide», «attrs»: {«text»: «MT020880.1», «term_id»: «1805599854», «term_text»: «MT020880.1»}} MT020880.1 были объединены и использованы для выравнивания. Мы запустили стандартный конвейер клеток 10X Genomics с объединенным геномом человека и SARS-CoV-2 для получения матриц подсчета для каждого из 4 условий роста.На каждое условие в среднем приходилось от 10 000 до 15 000 отсчетов на клетку или в среднем от 2 400 до 3600 уникальных генов, обнаруженных для каждого состояния. Подготовка библиотеки и секвенирование были выполнены Йельским центром анализа генома.
4.4. Количественная ОТ-ПЦР SARS-CoV-2
Вирусная РНК из лизатов клеток HBEC, инфицированных SARS-CoV-2, экстрагировалась с использованием TRIzol (Life Technologies) и очищалась с использованием Direct-zol RNA MiniPrep Plus в соответствии с инструкциями производителя (Zymo Research, Irvine , Калифорния, США).Был проведен двухэтапный синтез кДНК с 5 мкл РНК, случайным гексамером и обратной транскриптазой ImProm-II (Promega, Мэдисон, Висконсин, США). Анализ qPCR выполняли в двух экземплярах для каждого из образцов и стандартных кривых, построенных с использованием нуклеокапсидных (N1) специфических олигонуклеотидов SARS-CoV-2 от Integrated DNA Technologies (Coralville, IA, USA): Зонд: 5 ‘6FAM ACCCCGCATTACGTTTGGTGGACC-BHQ1 3’ ; Прямой праймер: 5 ’GACCCCAAAATCAGCGAAAT 3’; Обратный праймер: 5 ’TCTGGTTACTGCCAGTTGAATCTG 3’.Предел обнаружения составил 10 копий генома SARS-CoV-2/ мкл . Количество копий вируса определяли количественно с использованием контрольной плазмиды, которая содержит полный ген нуклеокапсида из SARS-CoV-2.
4.5. Проверка полиаденилированных транскриптов SARS-CoV-2
Клетки Huh7.5, выращенные в DMEM, содержащей 10 процентов FBS, были инфицированы 10 4 БОЕ SARS-CoV-2, и клеточные лизаты были собраны на 0, 1, 2 и 3 точки. dpi. Используя 0,3 мкг тотальной РНК, экстрагированной из ложного или зараженного SARS-CoV-2 Huh7.5 клеток в разные моменты времени, обратная транскрипция была выполнена с помощью oligo-d (T) 20 (ThermoFisher) и MarathonRT, высокопроизводительной RT, кодируемой интроном группы II. Очистку MarathonRT и реакции RT проводили, как описано ранее [ 40 ]. ПЦР (NEBNext Ultra II Q5®Master Mix, NEB, Ипсвич, Массачусетс, США) выполняли с генно-специфическим прямым праймером, сконструированным на 700 н. Выше видимой границы между телом генома SARS-CoV-2 и предполагаемым поли- Хвост. Oligo-d (T) 20 использовали в качестве обратного праймера.Цикл Touchdown PCR использовали для повышения специфичности реакции PCR. Продукты ОТ-ПЦР разделяли на 1,3% агарозном геле с лестницей (ДНК-лестница 100 п.н., плюс ДНК-лестница 1 т.п.н., Invitrogen). Ниже приведены олигонуклеотиды прямой ПЦР, использованные в этом эксперименте, который включает два положительных контроля.
Положение названия праймерав последовательности генома 5’-3 ’:
| F_Val_1 | 7700 | GAGAGACTTGTCACTACAGTTTAAA |
| F_Val _ 2 | 26650 | AATTTGCCTATGCCAACAGGA |
| F_Val _ (+) ve_1 | 28600 | AGATCTCAGTCCAAGATGGTA |
| F_Val _ (+) ve _ 2 | 29000 | GGTAAAGGCCAACAACAACAA |
4.6. Электронная микроскопия
Клетки фиксировали с использованием 2,5% глутарового альдегида в 0,1 М фосфатном буфере, осмифицировали в 1% тетроксиде осмия и дегидратировали в возрастающих концентрациях этанола. Во время дегидратации к 70% этанолу добавляли 1% уранилацетат для усиления ультраструктурного контраста мембраны. После обезвоживания клетки помещали в дуркупан. Ультратонкие срезы размером 70 нм вырезали на ультрамикротоме Leica, собирали на однослотовых решетках, покрытых Formvar, и анализировали с помощью электронного микроскопа Tecnai 12 Biotwin (FEI).
4.7. Измерение цитокинов с помощью мультиплексного иммуноанализа
Уровни IL6, IL1A, IL1B, IL1RN и CXCL9 в базолатеральных супернатантах ложных и инфицированных HBEC были выполнены с помощью технологий EVE (Калгари, AB, Канада) с использованием мультиплексного иммуноанализа, проанализированного с помощью BioPlex 200 Весь статистический анализ был выполнен с использованием Prism GraphPad версии 8. Все статистические анализы были проанализированы с использованием непараметрического критерия Краскела-Уоллиса, обозначены полосой, а значение p представлено символом.(* p-значение ¡0,05, ** p-значение ¡0,01, *** p ¡0,001).
4.8. Клинические образцы
Этическое заявление
Исследование на людях проводилось после одобрения Институционального наблюдательного совета Йельского университета и проводилось в соответствии с принципами, изложенными в Хельсинкской декларации. Письменное информированное согласие было получено от законного опекуна до взятия проб.
История болезни и сбор клинических образцов
У 15-летнего мужчины с ожирением, обструктивным апноэ во сне и метаболическим синдромом в течение 6 дней наблюдалась лихорадка (107 ° F ), головная боль, кашель, ощущение стеснения / боли в груди, усталость , и анорексия.Затем за 2 дня до госпитализации у пациента развилась одышка, ему потребовалась усиленная респираторная поддержка, и на 2-й день его курса лечения в больнице он был интубирован. Ему также поставили диагноз гипоксической дыхательной недостаточности и острого респираторного дистресс-синдрома (ОРДС), вторичных по отношению к инфекции COVID-19. Пациент оставался интубированным в течение 8 дней. Во время интубации у него был индекс оксигенации 28, и его рентгенограмма показала многоочаговые двусторонние помутнения воздушного пространства. Лабораторные химические исследования показали лимфопению с абсолютным числом лимфоцитов 0.5 × 1000/ мкл , повышенный уровень ферритина (2386 нг / мл), повышенный уровень d-димера (30,7 мг / л), повышенный уровень СРБ (298 мг / л), трансаминит (ALT 245 U / L, AST 119 U / L) ), повышенный BNP (835 пг / мл), повышенный IL-6 (37 пг / мл), слегка повышенный IL-10 (20 пг / мл) и слегка повышенный IL-2R (1203 пг / мл). На пятый день нахождения на ИВЛ у пациента развилась вторичная бактериальная пневномия, лечили антибиотиками. Контрольная пациентка — 15-летняя женщина, которой в начале 2019 года была интубирована плановая ортопедическая операция.Она прибыла в педиатрическое отделение интенсивной терапии интубированной для купирования боли и не имела никаких заболеваний легких. Она была экстубирована через несколько часов после прибытия, как только был установлен режим обезболивания. Эндотрахеальный аспират собирали во время интубации и экстубации. Было получено приблизительно 5 мл эндотрахеального аспирата и помещено на лед. Эндотрахеальный аспират пропускали через нейлоновый фильтр 100 мкм для удаления мусора и комков, центрифугировали и ресуспендировали в холодной среде DMEM с 10
4.9. Анализ данных scRNA-seq
4.9.1. Аннотация типа клетки
Мы использовали стандартный конвейер анализа scRNA-seq для кластеризации [ 41 ]. Вкратце, чтобы учесть выпадение транскриптов, присущее scRNA-seq, мы удалили гены, которые экспрессировались менее чем в 3 клетках, и удалили клетки, экспрессирующие менее 200 генов. Затем мы отфильтровываем клетки с более чем 10 процентами митохондриальных генов. Мы не обнаружили корреляции между числом копий вируса и экспрессией митохондрий.Полученные в результате подсчеты необработанных уникальных молекулярных идентификаторов (UMI) в каждой ячейке были нормализованы по размеру их библиотеки. Затем нормализованные подсчеты были преобразованы в квадратный корень, что аналогично логарифмическому преобразованию, но не требует добавления псевдосчетчика. Предварительная обработка данных выполнялась на Python (версия 3.7.4) с использованием Scanpy (версия 1.4.6) [ 42 ].
Мы визуально наблюдали пакетные эффекты между условиями в двумерных клеточных вложениях. Чтобы удалить эти пакетные эффекты для кластеризации, аннотации типов ячеек и визуализации, мы использовали приблизительный пакетно-сбалансированный график kNN для многообразного обучения (коррекция пакетного эффекта BB-kNN) с помощью реализации быстрого приближения Scanpy [ 43 , 42 ].BB-kNN предполагает, что по крайней мере некоторые типы ячеек являются общими для разных партий и что различия между партиями для одного и того же типа ячеек меньше, чем различия между ячейками разных типов в партии. Для каждой ячейки 3 ближайших соседних ячейки в каждом состоянии были идентифицированы евклидовым расстоянием в 100-мерном пространстве анализа главных компонентов (PCA). Этот график kNN был использован в качестве основы для последующего анализа.
Для визуализации данных scRNA-seq мы реализовали различные методы нелинейного уменьшения размеров и использовали матрицу связности BB-kNN с пакетной коррекцией в качестве входных данных для Uniform Manifold Approximation and Projection (UMAP) [ 44 ] и Potential of Heat- диффузия для построения траектории на основе аффинности (PHATE) [ 45 ].Проекции UMAP были созданы с использованием минимального расстояния 0,5. Проекции PHATE были созданы с параметром гамма 1.
Для кластеризации клеток мы использовали метод обнаружения сообщества Лувена [ 46 ] с графиком BB-kNN. Мы использовали обнаружение сообществ с высоким разрешением и объединенные кластеры на основе экспрессии маркеров типа клеток бронхиального эпителия, чтобы выделить некоторые редкие типы клеток, например Клетки пучка [ 47 , 48 ]. Чтобы аннотировать различные типы клеток, присутствующие в HBEC, мы проанализировали экспрессию ряда маркерных генов, о которых сообщалось в атласе молекулярных клеток от Travaglini et al.[ 47 ]. Мы сосредоточились на 8 типах клеток: (i) базальные клетки (KRT5, DAPL1, TP63), (ii) ресничные клетки (FOXJ1, CCDC153, CCDC113, MLF1, LZTFL1), (iii) клубные клетки (SCGB1A1, KRT15, CYP2F2, LYPD2 , CBR2), (iv) BC / клуб (KRT4, KRT13), (v) Нейроэндокринные клетки (CHG1, ASCL1), (vi) Клетки пучка (POU2F3, AVIL, GNAT3, TRPM5), (vi) Ионоциты (FOXI1, CFTR , ASCL3) и (viii) бокаловидные клетки (MUC5AC, MUC5B, GP2, SPDEF).
Чтобы проанализировать динамику сплайсинга в масштабе ~ часов и транскриптомную регуляцию внутри инфицированных клеток и клеток-свидетелей, претерпевающих изменения в течение нескольких дней воздействия SARS-CoV-2, мы выровняли показания для разделения сплайсированных и несплайсированных счетчиков [ 49 ].Из этих многоуровневых матриц подсчета мы вычисляем скорость РНК для отдельных клеток с помощью scVelo, обобщения исходного метода скорости РНК, который позволяет охарактеризовать множественные состояния транскрипции [ 27 , 49 ]. Мы вычисляем моменты первого и второго порядка с 50 ПК и предполагаем 30 соседей. Затем мы подобрали три динамические скоростные модели РНК, чтобы избежать искажающего смещения пакетных эффектов и узнать отдельные латентные времена из вычисленных скоростей РНК: одна модель, включающая все HBEC, и по одной модели для инфицированных клеток и клеток-свидетелей.С помощью scVelo мы визуализируем изученную динамику и взаимосвязь подсчетов сращенных и несвязанных, а также глобально просматриваем скорость в нашем исходном встраивании UMAP.
4.9.2. Порог инфицирования и показатель инфицирования
Подсчет вирусного транскрипта в клетке не означает, что клетка инфицирована, так как этот подсчет может происходить от вируса, прикрепленного к поверхности клетки, вируса окружающей среды в суспензии или из-за несовпадения считывания. Учитывая сообщенный общий 3 ’поли (A) хвост в транскриптах коронавируса [ 50 ], мы не были уверены, сможем ли мы правильно захватить различные ORF с помощью библиотеки экспрессии гена 10x Genomics 3’.Таким образом, мы выровняли вирусные считывания с одним «экзоном» всего генома, то есть подсчитали считывание, сопоставленное с ORF SARS-CoV-2 и межгенными областями. Эти подсчеты использовались для определения инфекционного состояния отдельных клеток. Чтобы отфильтровать клетки с количеством транскриптов вирусного генома, которое может быть результатом прикрепления вирусных клеток к поверхности, окружающего вируса в суспензии капель или несоответствия считывания, мы считали инфицированные клетки имеющими ≥ 10 счетчиков вирусных транскриптов. Это значение было определено порогом количества вирусов в имитационном состоянии.Несмотря на то, что в фиктивном состоянии не ожидается количества вирусов, мы наблюдали небольшое количество, которое мы относим к несовпадению. Мы наблюдали только 5 фиктивных клеток с полным числом транскриптов вирусного генома SARS-CoV-2 ≥ 10 транскриптов. Эти критерии позволили нам найти 144 инфицированных клетки при 1 dpi, 1428 ячеек при 2 dpi и 3173 клетки при 3 dpi.
Для количественной оценки степени, в которой отдельная клетка транскрипционно похожа на инфицированную клетку, мы использовали ранее разработанный подход обработки графического сигнала, названный Manifold Enhancement of Latent Dimensions (MELD) [ 51 ].Мы закодировали необработанную экспериментальную оценку для каждой ячейки в наборе данных таким образом, что -1 представляет наблюдателя или незараженную ячейку, а +1 представляет инфицированную ячейку. Используя ядро из графика BB-kNN (описанного выше), эти необработанные оценки были сглажены в области графика, давая «балл инфекции» для каждой клетки, который представляет степень, в которой отдельная клетка транскрипционно подобна инфицированным клеткам. Для сводной статистики этот балл стратифицирован по типу и состоянию клеток.
4.9.3. Анализ охвата считыванием вирусного генома
Для визуализации охвата считыванием вируса вдоль вирусного генома мы использовали файлы двоичной карты выравнивания (BAM) со штрих-кодом (BAM) 10X Genomics для каждого образца. Мы отфильтровали файлы BAM, чтобы сохранить только сопоставление считываний с вирусным геномом, используя инструмент , пересекающий [ 52 ]. Мы преобразовали файлы BAM в файлы карты выравнивания последовательностей (SAM), чтобы отфильтровать ячейки, которые были удалены в нашем конвейере предварительной обработки данных отдельных ячеек.Глубина секвенирования для каждой базовой позиции была рассчитана с использованием счетчика образцов . Чтобы охарактеризовать распределение чтения по вирусному геному, мы подсчитали транскрипты 10 различных ORF: ORF1ab, поверхностный гликопротеин (S), ORF3a, белок оболочки (E), мембранный гликопротеин (M), ORF6, ORF7a, ORF8, фосфопротеин нуклеокапсида (N) и ORF10.
4.9.4. Анализ дифференциальной экспрессии
Чтобы найти дифференциально экспрессируемые гены в разных условиях, мы использовали комбинацию трех показателей: расстояние Вассерштейна или земного движителя, скорректированное значение p из двустороннего U-критерия Манна-Уитни с непрерывностью и поправкой Бенджамини-Хохберга. , и двоичный логарифм кратного изменения между средними значениями.Значимость была установлена на p скорректирована ≤ 0,01. Расстояние Земного Двигателя, или одномерное расстояние Вассерштейна, можно определить как минимальную стоимость преобразования одного распределения в другое, и оно ранее использовалось для оценки экспрессии генов, которые существенно различаются между условиями [ 53 , 54 ]. Мы выполнили несколько бинарных сравнений для каждой временной точки и для объединения 1, 2 и 3 точек на дюйм: инфицированные против сторонних наблюдателей, зараженные против.фиктивные ячейки, и сторонний наблюдатель против фиктивных ячеек. На тепловых картах были представлены 30 генов с наиболее дифференцированной экспрессией (с повышенной или пониженной регуляцией, ранжированные по расстоянию Вассерштейна) в каждом состоянии, типе клеток и анализе. Чтобы идентифицировать предполагаемые клеточные функции, изменяющиеся в разных условиях, мы выполнили тесты статистической избыточной репрезентативности PANTHER-GO [ 55 ] для активируемых генов в каждом типе клеток и каждом состоянии отдельно, используя стандартный справочный список Human PANTHER-GO в качестве фона.
4.10. Встраивание условной плотности (CONDENSE) для объективной характеристики динамики генов
Чтобы охарактеризовать гены на основе их временной динамики, мы разработали новый вычислительный метод, который сочетает в себе идеи обработки графовых сигналов, оценки плотности, теории информации и множественного обучения. Наш метод называется CON ditional DENS год E mbedding (или CONDENSE) предназначен для вывода временной динамики, оценки совместной и условной плотности, а затем встраивания генов с использованием этих плотностей.показывает обзор метода CONDENSE. Цель CONDENSE состоит в том, чтобы получить встраивание генов в соответствии с их динамикой транскрипции во времени, чтобы это вложение можно было использовать для обнаружения новых сигнатур динамики генов.
4.10.1. Обзор и обозначение CONDENSE
Алгоритм CONDENSE для анализа динамики генов в данных отдельных клеток выглядит следующим образом:
Получите интересующее условие с помощью псевдовременного или другого метода вывода траектории
Получите общую оценку плотности по клеткам для каждого гена как функцию времени с помощью оценки плотности ядра
Нормализовать оценку плотности для каждого гена по его временному условию для получения условной плотности встраивания генов
Используйте встраивание динамики генов для последующего анализа, такого как визуализация, кластеризация, архетипический анализ и обогащение набора генов
Хотя метод был разработан с использованием одного Имея в виду данные выражения ячеек, CONDENSE может применяться к любому набору данных из m примеров X = {x (1),…, x (m)} с условием c .Здесь мы рассматриваем каждый ген как образец с экспрессией этого гена в наборе данных scRNA-seq, представленным вектором x ( i ) для конкретного гена i . Мы выводим псевдовремя для представления условия c . Наконец, чтобы сравнить дифференциальную динамику генов между интересующими нас слоями, S k для k ∈ {1, 2 ,… l }, мы вычисляем условное вложение плотности для каждого слоя отдельно, а затем объединяем оценки для получения совместного распределения,
pm (X∣c) = pm, S1 ~ X (XS1∣cS1) pm, S2 ~ X (XS2∣cS2) ‖… ‖pm, Sk ~ X (XSk∣cSk)
(1)
где pm (X∣c) — это условная плотность встраивания, представленная в виде вектора, для гена m .Для всех генов мы получаем матрицу встраивания условной плотности PSk для k слоев, которая имеет условно нормализованную оценку совместной плотности вероятности для m генов. Из этого набора условно нормализованных совместных распределений вероятностей мы выполняем многообразное обучение для неконтролируемого представления динамики генов. Сопоставляя динамику генов для двух интересующих нас слоев, а именно, инфицированных и случайных клеток, мы эффективно вычисляем расстояние Вассерштейна (или расстояние земного движителя, EMD) между условно нормализованными совместными распределениями вероятностей, EMD (pSinfected ~ X, pSbystander ~ X) [ 56 ].Далее мы подробно опишем каждый аспект CONDENSE.
4.10.2. Вывод псевдовременного условия
Чтобы вывести временное состояние c на основе отдельной клетки, ограниченное транскриптомом конкретной клетки, мы использовали псевдовременной вывод или вывод о траектории. Существует несколько методов вывода псевдодействия; однако не всегда ясно, какой базовый динамический процесс фиксируется латентным или псевдовременным представлением [ 57 ]. Чтобы гарантировать, что наше состояние соответствует динамике в течение нескольких дней после заражения, а не другим формам клеточной гетерогенности, мы использовали контролируемый подход, чтобы вывести псевдовремя, сглаживая известную метку временной точки клетки на континуум в соответствии с транскциптомным сходством между клетками. с использованием Manifold Enhancement of Latent Dimensions (MELD) [ 51 ].В нашей реализации мы использовали α -распадающееся ядро с расстояниями и связностями, полученными из приближенного графа BB-kNN, что позволило нам определить матрицу подобия,
Wi, j = 12 (e− (‖xi − xj‖2ϵk (xi)) α + e− (‖xi − xj‖2ϵk (xj)) α)
(2)
где ϵ k ( x i ) и ϵ k ( x j ) — евклидовы расстояния в пространстве PCA между ячейками x i и x j для k соседей i с ненулевыми связями.Мы можем получить полную вариацию данного сигнала f∈ℝn для нашего графа подобия W по n вершинам через лапласовскую квадратичную форму,
fTLf = ∑i, jWi, j (f (i) −f (j)) 2
(3)
где L — лапласиан графа, а f ( i ) — метка временной точки для данная пара ячеек и . Здесь мы назначаем f для данной ячейки i как f i = t i для t ∈ {0, 1, 2, 3}, указывая, Ячейка была получена из образца Mock, 1, 2 или 3 dpi.(λ) = [I + βL − αI) ρ] −1
(4)
где λ — собственные значения Ψ , — параметр модуляции (установлен на 40), β — штраф восстановления ( установлен в 1) и определяет порядок фильтра (установлен в 2). Чтобы получить псевдовремя, мы следуем реализации [ 51 ], которая вычисляет частотную характеристику или сглаженный сигнал, используя приближение полинома Чебышева, чтобы получить H [ 58 ].
4.10.3. Оценка условной плотности
Далее мы вычисляем оценку условной плотности для каждого слоя и экспрессии каждого гена как функцию псевдовремени.Em, k (xm) = 1hNk∑i∀i∈kK (xm − xi, mh)
(5)
, где h — полоса пропускания ядра K , p Em, k — это плотность вероятности для k -ого слоя клеток в 95 процентильном диапазоне экспрессии в этом слое, N k — это количество клеток в этом слое и x i , m — нормализованные значения для гена m в ячейке i . m, k (x = z) = pEm, k (x = z) ⋅1hn⋅∑z = 1nptm, k (t = z ′)
(6)
где x = z задает оценку плотности для ячеек в интервале бинирования z , а t = z ‘задает ячейки в бинтовом состоянии или псевдовременном интервале z ‘.Здесь мы предполагаем, что z и z ′ являются интервалами фиксированной ширины, так что z ( n ′) = z ( n ′ −1) + Δ z где Δz = hn (max x − min x) и z ′ ( n ′) = z ′ ( n ′ −1) + Δ z , где Δz = hn (max t − min t), где n ′ — параметр индексации ∈ 1, 2,… l до n и на практике мы ограничиваем диапазон x и t 95-м процентильный диапазон.Таким образом, мы получаем оценку условной плотности для каждого гена и страты после итерации по каждому выражению с разбивкой на интервалы и условному интервалу.
4.10.4. Вложение с условной плотностью
На практике уравнение 6 можно быстро вычислить, используя двумерную гистограмму экспрессии гена в зависимости от псевдовременности и нормализуя каждый интервал на сумму по распределению экспрессии генов за интервал псевдовременного бина. Здесь мы используем 20 равномерно расположенных интервалов в диапазоне 95–-го процентилей экспрессии генов и псевдодействия.Мы повторяем этот процесс для каждого гена и сглаживаем условно нормализованную двумерную гистограмму (оценка условной плотности) P m , k в вектор p m , k . Затем мы объединяем векторы для каждого слоя в соответствии с уравнением 1, чтобы получить матрицу условных оценок плотности по m генам , PSk∈ℝm × (c × k), где m -я строка и c — -й столбец представляет собой условную оценку плотности для гена m в псевдовремя c в конкретном интересующем пласте, повторенную k раз.Таким образом, мы получаем нашу условную плотность вложения по интересующим пластам к , PSk.
4.10.5. Архетипический анализ матрицы CONDENSE
Чтобы найти экстремальную динамику на основе отдельного гена между слоями, мы выполнили архетипический анализ на матрице оценки условной плотности, PSk. Архетипический анализ оценивает основную выпуклую оболочку набора данных d -симплекс [ 59 ]. Выпуклый корпус размером d можно рассматривать как оболочку вокруг данных.Мы реализуем архетипический анализ, определяя главную выпуклую оболочку как ( d — 1) -мерную выпуклую оболочку, которая наилучшим образом учитывает данные в соответствии с
arg min Q, YD (PSk∣PSkQY)
(7)
где D — мера искажения, а Q и Y — оптимальные матрицы, которые минимизируют искажение D с учетом ограничений, | q d | 1 = 1 и Q ≥ 0, а | л м | 1 = 1 и Y ≥ 0 соответственно.Эти ограничения создают архетипы,
, которые представляют собой выпуклые комбинации, которые могут быть аппроксимированы средневзвешенным значением векторов признаков PSk∈ℝcxd [ 60 ]. В этой реализации анализа основной выпуклой оболочки (PCHA) мы идентифицируем архетипы A , которые предпочитают особенности, представляющие «углы» данных, что позволяет идентифицировать архетипы или отдельные и экстремальные аспекты данных [ 60 ] . Чтобы оценить d -мерность наших данных оценки условной плотности, мы использовали «метод локтя» кумулятивной дисперсии, объясненной главными компонентами, после вычисления PCA (PSk) для получения ( d — 1) архетипов.Архетипический анализ — это один из многих методов аппроксимации матриц низкого ранга, одним из которых является более широко используемый анализ главных компонент.
4.10.6. Биологические выводы из CONDENSE
После определения динамики отдельных генов с помощью архетипического анализа, мы дополнительно охарактеризуем набор генов, выраженных в нашем наборе данных, путем визуализации многомерных, условных оценок плотности в одномерном сокращенном вложении. Возможен любой метод обучения многообразию или метод уменьшения размерности, но мы используем популярный алгоритм UMAP для изучения отображения u: ℝc × k↦ℝ2 [ 44 ].Мы подбираем эту функцию для вложения условной плотности PSk и преобразуем PSk, чтобы получить вложение условной плотности PSk ′ в пространство UMAP. Чтобы охарактеризовать динамику в соответствии с архетипической или экстремальной динамикой в наборе данных HBEC, мы относим каждый ген к архетипу, минимизируя евклидово расстояние от точки включения условной плотности каждого гена до ближайшей архетипической точки в пространстве UMAP,
ai = arg min (AT) ′ ‖PSk ′ — (AT) ′ ‖22
(9)
где a i представляет собой архетип, удовлетворяющий задаче минимизации, ( A T ) представляет архетипы в координатах UMAP, а PSk ‘обозначает условную плотность встраивания генов на страты k в координатах UMAP.Это позволяет нам идентифицировать гены с четкой и дифференцированной динамикой в инфицированных клетках по сравнению с клетками-свидетелями в ходе инфекции и глобально визуализировать вариации в динамике генов при разрешении одного гена.
Мы внутренне проверяем CONDENSE с помощью локального сглаживания диаграммы рассеяния (LOESS) [ 61 ]. Вкратце, мы визуализируем экспрессию генов для интересных архетипов на основе предполагаемого псевдовремени клеток, чтобы подтвердить, что оценка условной плотности соответствует динамике псевдовремени для каждого слоя.Это также обеспечивает дополнительный способ оценки дифференциальной динамики между слоями, хотя и при условии, что изменения в экспрессии генов локально линейны по псевдовремени. После получения CONDENSE можно провести множество последующих анализов, в том числе с использованием связывания архетипов и генов с другими метаданными клеток или генов посредством обучения с учителем. Однако, чтобы обеспечить дополнительное понимание генов, демонстрирующих отличную и дифференциальную динамику между инфицированными и случайными клетками, мы выполнили анализ PANTHER-GO на генах в различных архетипических кластерах и идентифицировали избыточно представленные пути в архетипах, демонстрирующих интересную динамику.