
Сайт vpr-klass.com — впр-класс.ком : гдз, решебник, гиа, егэ, решение задач, задания, варианты, подготовка к экзамену, тесты, презентации.
Error in links file
Сайт vpr-klass.com — впр-класс.ком : гдз, решебник, гиа, егэ, решение задач, задания, варианты, подготовка к экзамену, тесты, презентации. У нас вы найдете много учебных материалов: решебники, ГДЗ, тестовые задания, видео уроки, генераторы задач, решения упражнений гиа и егэ.
| Расскажи друзьям vpr-klass или впр-класс Презентации Детские презентации Презентации по математике Демо-варианты: Математика Русский язык Физика Обществознание Английский язык Информатика История Биология Химия Литература География Математика Русский язык |  com (впр-класс) com (впр-класс) Последние новости ГИА и ЕГЭ 2017. ЕГЭ по математике. КДР по математике. Математика 1-4 класс. Математика 5-6 класс. Алгебра и геометрия 7-9 класс. Алгебра и геометрия 10-11 класс. ГДЗ, решебники по математике, алгебре, геометрии. Онлайн калькуляторы по математике. Генераторы случайных примеров и задач по математике. Презентации. Другие школьные предметы. Сайт Vpr-klass.com — это учебный-образовательно-познавательный сайт для школьников! Приветствуем на уникальном сайте помощи всем ученикам 1-11 классов. На образовательном ресурсе полно полезной, учебной информации от способов решения заданий по математике до разных генераторов задач по алгебре и онлайн калькуляторов по геометрии, которые облегчат жизнь школьника. В частности, сделан больший уклон на решебники и ГДЗ, ведь правильная домашняя работа — это хорошие оценки и учеба в школе. | Интересно Много разных решений Тесты ГИА онлайн. Видео — ГИА 2013: геометрия Видео — ГИА 2012 Видео — Демо-вариант 2012. Решение Демо-варианта 2013 года (2014 года). Задача №1, Вычислить. Задача №2, Числа и прямая. Задача №3, Сравнение чисел. Задача №4, Уравнения. Задача №5, Графики и формулы. Задача №6, Прогрессии. Задача №8, Неравенства, системы неравенств. Задача №9, Задания по геометрии. Генератор вариантов ГИА 2014 Много разных решений. Онлайн тесты. Видео уроки ЕГЭ по математике. Генератор вариантов ЕГЭ 2014 Книги, справочники Решение демо варианта ЕГЭ по математике 2014 Задания B1, задача. Задания B2, диаграммы. Задания B5, уравнения. Задания B8, производная. Задания B10, вероятность. Видео уроки |



Copyright © 2017 vpr-klass.com | Если какой-либо из материалов нарушает ваши авторские права, просим немедленно связаться с Администрацией!!! Наш e-mail: [email protected] | Правообладателям |
sitemap. xml
xml
ГДЗ Математика за 5 класс Истер О.С.
Классы
1 2 3 4 5 6 7 8 9 10 11Математика
1 2 3 4 5 6 7 8 9 10 11 Английский язык
1
2
3
4
5
6
7
8
9
10
11 Русский язык
1
2
3
4
5
6
7
8
9
10
11Алгебра
1 2 3 4 5 6 7 8 9 10 11 Геометрия
1
2
3
4
5
6
7
8
9
10
11Физика
1 2 3 4 5 6 7 8 9 10 11Химия
1 2 3 4 5 6 7 8 9 10 11Немецкий язык
1 2 3 4 5 6 7 8 9 10 11Белорусский язык
1 2 3 4 5 6 7 8 9 10 11Украинский язык
1 2 3 4 5 6 7 8 9 10 11Французский язык
1 2 3 4 5 6 7 8 9 10 11Биология
1 2 3 4 5 6 7 8 9 10 11История
1 2 3 4 5 6 7 8 9 10 11Информатика
1 2 3 4 5 6 7 8 9 10 11ОБЖ
1 2 3 4 5 6 7 8 9 10 11География
1 2 3 4 5 6 7 8 9 10 11Природоведение
1 2 3 4 5 6 7 8 9 10 11Основы здоровья
1 2 3 4 5 6 7 8 9 10 11Музыка
1 2 3 4 5 6 7 8 9 10 11Литература
1 2 3 4 5 6 7 8 9 10 11Обществознание
1 2 3 4 5 6 7 8 9 10 11Черчение
1 2 3 4 5 6 7 8 9 10 11Мед.
 подготовка 1
2
3
4
5
6
7
8
9
10
11
подготовка 1
2
3
4
5
6
7
8
9
10
11Окружающий мир
1 2 3 4 5 6 7 8 9 10 11Человек и мир
1 2 3 4 5 6 7 8 9 10 11Астрономия
1 2 3 4 5 6 7 8 9 10 11Экология
1 2 3 4 5 6 7 8 9 10 11Технология
1 2 3 4 5 6 7 8 9 10 11Естествознание
1 2 3 4 5 6 7 8 9 10 11Испанский язык
1 2 3 4 5 6 7 8 9 10 11Искусство
1 2 3 4 5 6 7 8 9 10 11Китайский язык
1 2 3 4 5 6 7 8 9 10 11Кубановедение
1 2 3 4 5 6 7 8 9 10 11Казахский язык
1 2 3 4 5 6 7 8 9 10 11Мир природы и человека
1 2 3 4 5 6 7 8 9 10 11Классы
1 2 3 4 5 6 7 8 9 10 11ГДЗ вправа 681 математика 5 класс Истер – Telegraph
➡➡➡ ПОДРОБНЕЕ ЖМИТЕ ЗДЕСЬ!
ГДЗ вправа 681 математика 5 класс Истер
Подробный решебник (ГДЗ ) по Математике за 5 (пятый ) класс — готовый ответ вправа — 681 . Авторы учебника: Истер . Автор: Истер О .С . Издательство: Генеза год .
Авторы учебника: Истер . Автор: Истер О .С . Издательство: Генеза год .
Подробное решение вправа № 681 по математике для учащихся 5 класса , авторов Истер . ГДЗ по математике 5 класс Истер вправа — 681 .
Решебник (ГДЗ ) Математика 5 клас Істер 2019 ❤ . Відповіді до підручника нова програма ❤, ответы к учебнику онлайн на телефоне . .
ГДЗ математика 5 клас — (решебники) за підручником автора Істер, відео розв’язання . До вашої уваги всі номери до ГДЗ математика 5 клас Істер . Для користувачів нашого сайту готові домашні завдання існують усюди, але лише у нас ви отримуєте велику зручність у користуванні, також у . .
ГДЗ (готові домашні завдання): Математика 5 клас .О . С . Істер . Відповіді до підручника . Зручно користуватися з Ваших стфонів! Математика – дуже цікава і корисна наука . Вона знадобиться в житті кожному: і бізнесмену, і будівельнику, і кухарю, і айтішнику .
Відповіді та розв’язки вправ з математики . Вправа 681 математика Істер 2019 гдз 5 клас .
Ученики пятого класса в постоянном режиме сотрудничают со справочником по математике . Книга-помощница нужна ребятам для достижения Школьники любят наш портал Vshkole . Это обосновано тем, что у нас сотрудничать с ГДЗ по математике 5 класс Истер легко и комфортно .
На сайте собраны все готовые домашние задания школьной программы .
Готові домашні завдання (гдз ) з математики для 5 класу, автор Істер О .С . 2019 рік . Нова програма . Готові домашні завдання з математики 5 клас Істера О .С . (Нова програма, 2019) можуть допомогти учням в освоєнні обов’язкового рівня математичної підготовки .
ГДЗ (відповіді) Математика 5 клас Істер 2019 . Posted on 28 .09 .2020 by ukrdz . Кількість сторінок: 288 . Формат підручника: PDF . Дивитись онлайн та читати відповіді до підручника Математика 5 клас О .С . Істер – Нова програма 2019 рік .
Математика / 5 класс / Істер Решебник .
ГДЗ по математике для 5 класса — Герасимов . Авторы . Герасимов В . Д ., Пирютко О . Н ., Лобанов А . П . Издание подготовлено группой педагогов, которые постарались облегчить участь учащихся 5 класса , часами бьющихся над «домашкой» по математике .
Н ., Лобанов А . П . Издание подготовлено группой педагогов, которые постарались облегчить участь учащихся 5 класса , часами бьющихся над «домашкой» по математике .
Розв’язання завдань до підручника 5 клас Математика Істер О ., Генеза, 2019 рік< . .
Варто зауважити, що у нас ГДЗ з математики 5 клас Істер є завжди відкритим . Воно безкоштовне, тому батькам не потрібно витрачати фінанси на придбання своєму чаду книги . Також, доступ до файлу у нас вільний цілодобово .
ТУТ є повні та якісні відповіді ГДЗ Математика 5 клас автор Істер О .С . 2019 Генеза онлайн Нова програма Вправа номер задача Самостійна робота . .
Подробный решебник (ГДЗ ) по Математике за 5 (пятый ) класс — готовый ответ вправа — 681 . Авторы учебника: Истер . Автор: Истер О .С . Издательство: Генеза год .
Подробное решение вправа № 681 по математике для учащихся 5 класса , авторов Истер . ГДЗ по математике 5 класс Истер вправа — 681 .
Решебник (ГДЗ ) Математика 5 клас Істер 2019 ❤ . Відповіді до підручника нова програма ❤, ответы к учебнику онлайн на телефоне . .
Відповіді до підручника нова програма ❤, ответы к учебнику онлайн на телефоне . .
ГДЗ математика 5 клас — (решебники) за підручником автора Істер, відео розв’язання . До вашої уваги всі номери до ГДЗ математика 5 клас Істер . Для користувачів нашого сайту готові домашні завдання існують усюди, але лише у нас ви отримуєте велику зручність у користуванні, також у . .
ГДЗ (готові домашні завдання): Математика 5 клас .О . С . Істер . Відповіді до підручника . Зручно користуватися з Ваших стфонів! Математика – дуже цікава і корисна наука . Вона знадобиться в житті кожному: і бізнесмену, і будівельнику, і кухарю, і айтішнику .
Відповіді та розв’язки вправ з математики . Вправа 681 математика Істер 2019 гдз 5 клас .
Ученики пятого класса в постоянном режиме сотрудничают со справочником по математике . Книга-помощница нужна ребятам для достижения Школьники любят наш портал Vshkole . Это обосновано тем, что у нас сотрудничать с ГДЗ по математике 5 класс Истер легко и комфортно .
На сайте собраны все готовые домашние задания школьной программы .
Готові домашні завдання (гдз ) з математики для 5 класу, автор Істер О .С . 2019 рік . Нова програма . Готові домашні завдання з математики 5 клас Істера О .С . (Нова програма, 2019) можуть допомогти учням в освоєнні обов’язкового рівня математичної підготовки .
ГДЗ (відповіді) Математика 5 клас Істер 2019 . Posted on 28 .09 .2020 by ukrdz . Кількість сторінок: 288 . Формат підручника: PDF . Дивитись онлайн та читати відповіді до підручника Математика 5 клас О .С . Істер – Нова програма 2019 рік .
Математика / 5 класс / Істер Решебник .
ГДЗ по математике для 5 класса — Герасимов . Авторы . Герасимов В . Д ., Пирютко О . Н ., Лобанов А . П . Издание подготовлено группой педагогов, которые постарались облегчить участь учащихся 5 класса , часами бьющихся над «домашкой» по математике .
Розв’язання завдань до підручника 5 клас Математика Істер О ., Генеза, 2019 рік< . .
Варто зауважити, що у нас ГДЗ з математики 5 клас Істер є завжди відкритим . Воно безкоштовне, тому батькам не потрібно витрачати фінанси на придбання своєму чаду книги . Також, доступ до файлу у нас вільний цілодобово .
Воно безкоштовне, тому батькам не потрібно витрачати фінанси на придбання своєму чаду книги . Також, доступ до файлу у нас вільний цілодобово .
ТУТ є повні та якісні відповіді ГДЗ Математика 5 клас автор Істер О .С . 2019 Генеза онлайн Нова програма Вправа номер задача Самостійна робота . .
ГДЗ глава 20 / § 20.11 14 химия 9 класс Гузей, Сорокин
ГДЗ вопросы 23 физика 7 класс Перышкин
ГДЗ часть 1. страница 30 английский язык 3 класс рабочая тетрадь Happy English Кауфман, Кауфман
ГДЗ страница 95 английский язык 9 класс новый курс (5-ый год обучения) Афанасьева, Михеева
ГДЗ § 2 33 геометрия 10‐11 класс Погорелов
ГДЗ упражнения 130 алгебра 9 класс Макарычев, Миндюк
ГДЗ часть 3 (страница) 24 русский язык 4 класс рабочая тетрадь Соловейчик, Кузьменко
ГДЗ раздел 1 D английский язык 11 класс Контрольные (тестовые) задания Эванс, Дули
ГДЗ номер 1250 алгебра 7 класс Макарычев, Миндюк
ГДЗ задание 804 математика 5 класс Никольский, Потапов
ГДЗ § 21 10 алгебра 9 класс Задачник Мордкович
ГДЗ §13 11 обществознание 7 класс Боголюбов, Городецкая
ГДЗ упражнение 720 математика 6 класс Истомина
ГДЗ страница 206 английский язык 8 класс новый курс (4-ый год обучения) Афанасьева, Михеева
ГДЗ Часть 1 (упражнение) 9 литература 2 класс тетрадь для самостоятельной работы Малаховская
ГДЗ повторение 91 алгебра 9 класс Учебник, Задачник (2018) Мордкович, Семенов
ГДЗ страница 19 геометрия 8 класс контрольные работы Мельникова
ГДЗ самостоятельные работы / С-3 / вариант 1 4 математика 5 класс дидактические материалы Потапов, Шевкин
ГДЗ страница 11 английский язык 2 класс рабочая тетрадь с контрольными работами Enjoy English Биболетова, Денисенко
ГДЗ номер 1002 математика 5 класс Дорофеев, Шарыгин
ГДЗ параграф 21 обществознание 6 класс рабочая тетрадь Соболева
ГДЗ Глава первая. Углеводороды и их природные источники / § 8. Нефть и способы её переработки 9 химия 10 класс Габриелян
Углеводороды и их природные источники / § 8. Нефть и способы её переработки 9 химия 10 класс Габриелян
ГДЗ 10 класс / тема 10 / дополнительное задание 12 химия 10‐11 класс дидактический материал Радецкий
ГДЗ обучающие работы / О-29 9 алгебра 8 класс дидактические материалы Евстафьева, Карп
ГДЗ упражнение 173 русский язык 6 класс Практика Лидман-Орлова, Пименова
ГДЗ задание 310 математика 6 класс Никольский, Потапов
ГДЗ упражнение 905 алгебра 9 класс Мерзляк, Полонский
ГДЗ задание 527 математика 5 класс Никольский, Потапов
ГДЗ Глава вторая. Кислород- и азотсодержащие органические соединения и их природные источники / § 15. Дисахариды и полисахариды 7 химия 10 класс Габриелян
ГДЗ упражнение 666 русский язык 6 класс Львова, Львов
ГДЗ параграф 2 6 алгебра 7 класс рабочая тетрадь Миндюк, Шлыкова
ГДЗ упражнение 112 алгебра 7 класс Макарычев, Миндюк
ГДЗ номер 858 алгебра 7 класс Макарычев, Миндюк
ГДЗ упражнение 539 русский язык 6 класс Ладыженская, Баранов
ГДЗ вправа 365 украинский язык 7 класс Заболотный, Заболотный
ГДЗ упражнение 189 русский язык 6 класс Ладыженская, Баранов
ГДЗ страница 75 русский язык 1 класс рабочая тетрадь Тихомирова
ГДЗ вправа 559 алгебра 8 класс Кравчук, Пидручна
ГДЗ глава 3. задача 204 геометрия 7‐9 класс Атанасян, Бутузов
задача 204 геометрия 7‐9 класс Атанасян, Бутузов
ГДЗ самостоятельные работы / СР-54. вариант 2 физика 9 класс самостоятельные и контрольные работы Марон, Марон
ГДЗ страница 122 английский язык 11 класс Happy english Кауфман, Кауфман
ГДЗ текстовые задачи / уроки 1-6 8 математика 4 класс дидактические материалы Козлова, Гераськин
ГДЗ вариант 3 195 геометрия 8 класс дидактические материалы Мерзляк, Полонский
ГДЗ параграф 4-5 19 алгебра 9 класс рабочая тетрадь Ткачева, Федорова
ГДЗ С-3. вариант 1 алгебра 7 класс самостоятельные работы Александрова
ГДЗ № 539 математика 5 класс Зубарева, Мордкович
ГДЗ для тех, кому интересно / вариант 3 6 алгебра 7 класс дидактические материалы к учебнику Макарычева Звавич, Дяконова
ГДЗ упражнение 281 русский язык 7 класс Баранов, Ладыженская
ГДЗ номер 743 алгебра 7 класс Макарычев, Миндюк
ГДЗ страница 64 английский язык 8 класс рабочая тетрадь Starlight Баранова, Дули
ГДЗ номер 1203 алгебра 7 класс Макарычев, Миндюк
ГДЗ Виноградова 2
ГДЗ Страница 200
Гдз По Русскому 10 Класс Рыбченков
Гдз По Математике Вариант
Мутагенез богатого цистеином клип-домена в протеазе, формирующей паттерн дрозофилы, Snake
- Список журналов
- Рукописи авторов HHS
- PMC2518291
Arch Biochem Biophys. Авторская рукопись; доступно в PMC 200915 июля.
Авторская рукопись; доступно в PMC 200915 июля.
Опубликовано в окончательной редакции как:
Arch Biochem Biophys. 2008 г., 15 июля; 475(2): 169–174.
Published online 2008 May 1. doi: 10.1016/j.abb.2008.04.030
PMCID: PMC2518291
NIHMSID: NIHMS57196
PMID: 18477463
a, b and a, *
Информация об авторе Информация об авторских правах и лицензиях Отказ от ответственности
- Дополнительные материалы
Общим мотивом, обнаруженным в сериновых протеазах беспозвоночных, участвующих в иммунитете и развитии, является clip-домен, предположительно регулирующий каталитическую активность или белок-белковые взаимодействия в протеолитических каскадах. Змея функционирует в каскаде, который паттернирует эмбрион Drosophila и обеспечивает доступную модель для изучения структурных требований для функции клипового домена. Мы протестировали зимогены Snake, несущие мутации, заряженные на аланин, в clip-домене, на их способность спасать эмбрионы, лишенные эндогенного Snake, и на их взаимодействие посредством котрансфекции клеток S2 с вышерасположенными дефектами гаструляции и нижестоящими Easter в протеазном каскаде. Из 13 одиночных и множественных замен один двойной мутант в предполагаемой выступающей области проявлял серьезные дефекты при спасении эмбрионов, но обнаруживал только минимальные дефекты в анализе котрансфекции. Мы обсуждаем последствия этих и других результатов для потенциальной биологической роли домена клипа Snake и для использования анализа in vitro для предсказания поведения протеазы.
Мы протестировали зимогены Snake, несущие мутации, заряженные на аланин, в clip-домене, на их способность спасать эмбрионы, лишенные эндогенного Snake, и на их взаимодействие посредством котрансфекции клеток S2 с вышерасположенными дефектами гаструляции и нижестоящими Easter в протеазном каскаде. Из 13 одиночных и множественных замен один двойной мутант в предполагаемой выступающей области проявлял серьезные дефекты при спасении эмбрионов, но обнаруживал только минимальные дефекты в анализе котрансфекции. Мы обсуждаем последствия этих и других результатов для потенциальной биологической роли домена клипа Snake и для использования анализа in vitro для предсказания поведения протеазы.
Ключевые слова: сериновая протеаза, мутагенез заряженных в аланин, clip-домен, Drosophila , дорсовентральная полярность сериновые протеазы беспозвоночных и гомологи сериновых протеаз, обладающие регуляторной функцией в иммунитете или развитии [1]; например, 37 из 204 предсказанных родственных сериновым протеазам генов в Drosophila melanogaster содержат один или несколько clip-доменов [1, 2]. Его название было получено из наблюдения, что на рисунках, показывающих его дисульфидные связи, домен чем-то напоминает форму скрепки [3]. Клип-домены обычно состоят из 37–60 аминокислотных остатков, связанных между собой тремя строго консервативными дисульфидными связями, при этом остальные остатки имеют относительно низкое сходство [1, 2]. Предполагается, что эти домены играют роль в локализации протеаз, регуляции каталитической активности или в обеспечении взаимодействия с другими компонентами протеолитических каскадов [1, 4, 5], но для проверки этих гипотез было проведено относительно мало функциональных исследований.
Его название было получено из наблюдения, что на рисунках, показывающих его дисульфидные связи, домен чем-то напоминает форму скрепки [3]. Клип-домены обычно состоят из 37–60 аминокислотных остатков, связанных между собой тремя строго консервативными дисульфидными связями, при этом остальные остатки имеют относительно низкое сходство [1, 2]. Предполагается, что эти домены играют роль в локализации протеаз, регуляции каталитической активности или в обеспечении взаимодействия с другими компонентами протеолитических каскадов [1, 4, 5], но для проверки этих гипотез было проведено относительно мало функциональных исследований.
На сегодняшний день получена структурная информация только для трех клип-доменов, в том числе рентгеновская кристаллическая структура клип-домена из каталитически неактивного гомолога сериновой протеазы профенолоксидаза-активирующего фактора II (PPAF-II) из Holotrichia diomphalia [5 ] и структура двух клиповых доменов, полученная с помощью ЯМР-спектроскопии, в функциональной сериновой протеазе, активирующей профенолоксидазу, протеиназе-2 (PAP-2) из Manduca sexta [4]. Они представляют группу 3 и группу 2 из четырех подгрупп клип-доменов, определяемых гомологией последовательностей, где группы 1a (включая Drosophila Snake), 1б и 2 (в том числе Drosophila Easter) представляют собой clip-домены, ассоциированные с функциональными сериновыми протеазами, а группа 3 – ассоциированные с гомологами сериновых протеаз, не обладающими каталитической активностью [1, 2]. Структурные особенности, которые, вероятно, являются общими для всех клиповых доменов, включают трех-четырехцепочечный, нерегулярный β-лист, к которому петли, формирующие внешний край и дно заметной центральной щели, присоединены дисульфидными связями. Внешний край щели преимущественно состоит из гидрофильных остатков, полученных из последовательностей, лежащих между третьим и четвертым консервативными цистеинами (которые мы будем называть остатками петли 3), а внутренняя поверхность щели состоит из гидрофобных остатков. В PPAF-II ободок щели состоит из петель и короткой β-тяжи, тогда как в доменах clip PAP-2 этот ободок содержит две антипараллельные α-спирали, разделенные короткой соединительной петлей.
Они представляют группу 3 и группу 2 из четырех подгрупп клип-доменов, определяемых гомологией последовательностей, где группы 1a (включая Drosophila Snake), 1б и 2 (в том числе Drosophila Easter) представляют собой clip-домены, ассоциированные с функциональными сериновыми протеазами, а группа 3 – ассоциированные с гомологами сериновых протеаз, не обладающими каталитической активностью [1, 2]. Структурные особенности, которые, вероятно, являются общими для всех клиповых доменов, включают трех-четырехцепочечный, нерегулярный β-лист, к которому петли, формирующие внешний край и дно заметной центральной щели, присоединены дисульфидными связями. Внешний край щели преимущественно состоит из гидрофильных остатков, полученных из последовательностей, лежащих между третьим и четвертым консервативными цистеинами (которые мы будем называть остатками петли 3), а внутренняя поверхность щели состоит из гидрофобных остатков. В PPAF-II ободок щели состоит из петель и короткой β-тяжи, тогда как в доменах clip PAP-2 этот ободок содержит две антипараллельные α-спирали, разделенные короткой соединительной петлей. Также отмечается, что клип-домен PPAF-II имеет выступающую область или выступ, состоящий из остатков, лежащих между первым и вторым консервативными цистеинами (остатки петли 1).
Также отмечается, что клип-домен PPAF-II имеет выступающую область или выступ, состоящий из остатков, лежащих между первым и вторым консервативными цистеинами (остатки петли 1).
Мутация строго консервативного валина на дне центральной щели в PPAF-II отменяет связывание PPAF-II с частично процессированной профенолоксидазой, событие, необходимое для проявления фенолоксидазной активности [5]. Этот результат указывает на то, что центральная щель, законсервированная в clip-доменах, служит сайтом связывания для других белков, таких как субстраты. Потеря clip-домена из каталитически активной сериновой протеазы Истер-типа, PPAF-I, приводила к значительному снижению активации укороченного PPAF-I плазмой по сравнению с активацией интактного зимогена PPAF-I [5]. Эти авторы предположили, что clip-домены могут играть важную роль в связывании со стереотипными молекулярными структурами на поверхности патогенов, что может приводить многие молекулы clip-домена, содержащие протеазы в каскаде, в тесный контакт друг с другом и обеспечивать усиление сигнала в этом месте. Сходная роль в связывании патогенов была предложена для клип-доменов PAP-2, возможно, с участием положительно заряженного участка на β-слое, включающего остатки, лежащие в петле 1 или непосредственно на карбокси-конце второго цистеина [4]. Однако эти предполагаемые роли в поверхностном связывании не были проверены экспериментально.
Сходная роль в связывании патогенов была предложена для клип-доменов PAP-2, возможно, с участием положительно заряженного участка на β-слое, включающего остатки, лежащие в петле 1 или непосредственно на карбокси-конце второго цистеина [4]. Однако эти предполагаемые роли в поверхностном связывании не были проверены экспериментально.
Две сериновые протеазы, содержащие клип-домен, Snake и Easter, являются нижестоящими компонентами четырехчленного каскада сериновых протеаз, который устанавливает дорсовентральную ось эмбриона Drosophila посредством вентральной активации сигнального лиганда для рецептора Toll [6– 8]. Эти протеазы представляют собой привлекательную мишень для функциональных исследований клиповых доменов, поскольку доступен чувствительный анализ для проверки способности сконструированных in vitro транскрибируемых мРНК спасать эмбрионы, лишенные какой-либо из этих протеаз, при инъекции в первые часы эмбрионального развития [7, 9].]. Ранее было показано, что делеция clip-домена Snake или мутация любого из его консервативных цистеинов отменяет способность введенной мРНК змеи спасать змеиных -мутантных эмбрионов, демонстрируя важность clip-домена для функции Snake [9]. Кроме того, сообщалось, что транскрипты, кодирующие N-концевой продомен Snake или Easter, включая домен clip и другие последовательности, дорсализуют эмбрионы дикого типа, что позволяет предположить, что продомен может конкурировать за связывание с другими важными компонентами каскада, такими как гаструляция. Дефектная (GD) протеаза, активирующая змеиный субстрат, змеиный субстрат, истер или неохарактеризованные кофакторы [9].]. В текущей работе мы воспользовались анализом спасения эмбриона после инъекции и установленной моделью коэкспрессии дорсовентральных протеаз [10, 11] в клеточной культуре для мутации 10 заряженных остатков в домене клипа Snake на аланин, индивидуально и последовательно. групп для дальнейшего изучения структурных требований к домену клипа в функции Snake.
Кроме того, сообщалось, что транскрипты, кодирующие N-концевой продомен Snake или Easter, включая домен clip и другие последовательности, дорсализуют эмбрионы дикого типа, что позволяет предположить, что продомен может конкурировать за связывание с другими важными компонентами каскада, такими как гаструляция. Дефектная (GD) протеаза, активирующая змеиный субстрат, змеиный субстрат, истер или неохарактеризованные кофакторы [9].]. В текущей работе мы воспользовались анализом спасения эмбриона после инъекции и установленной моделью коэкспрессии дорсовентральных протеаз [10, 11] в клеточной культуре для мутации 10 заряженных остатков в домене клипа Snake на аланин, индивидуально и последовательно. групп для дальнейшего изучения структурных требований к домену клипа в функции Snake.
Сайт-направленный мутагенез кДНК змеи
Полноразмерная кДНК змеи , предоставленная Robert DeLotto [9] подвергали мутагенезу с использованием набора для направленного мутагенеза QuikChange и полимеразы PfuTurbo (Stratagene), а клоны секвенировали перед использованием в анализах. Праймеры, используемые для всех реакций мутагенеза, перечислены в дополнительной таблице 1. Последовательность клип-домена Snake (от F92 до P139) представляет собой FC RR SF D G R SGYCILAYQCLHVI RE Y R VHG48. I D ICTH R NNVPVICCP. Десять заряженных остатков (выделены жирным шрифтом) индивидуально мутировали в аланин, чтобы получить 10 отдельных мутантных конструкций. Два двойных мутанта, loop1 (R94A, D98A) и loop3R (R123A, D125A), а также один тройной мутант, loop3L (R115A, E116A, R118A), были созданы с помощью последовательных реакций мутагенеза.
Праймеры, используемые для всех реакций мутагенеза, перечислены в дополнительной таблице 1. Последовательность клип-домена Snake (от F92 до P139) представляет собой FC RR SF D G R SGYCILAYQCLHVI RE Y R VHG48. I D ICTH R NNVPVICCP. Десять заряженных остатков (выделены жирным шрифтом) индивидуально мутировали в аланин, чтобы получить 10 отдельных мутантных конструкций. Два двойных мутанта, loop1 (R94A, D98A) и loop3R (R123A, D125A), а также один тройной мутант, loop3L (R115A, E116A, R118A), были созданы с помощью последовательных реакций мутагенеза.
Функциональный анализ с помощью инъекции мРНК в эмбрионы
кДНК дикого типа и мутантной змеи в векторе pBS-SK- (Stratagene) линеаризовали с помощью Bam HI, разрезающего ниже змеи 3’UTR и полиА хвост, очищенный гелем и использованный в качестве матрицы для продукции in vitro полиаденилированной 5′-кэпированной мРНК с использованием набора T7 mMessage mMachine в соответствии с инструкциями производителя (Ambion). Экстрагированную фенолом-хлороформом, осажденную изопропанолом мРНК растворяли в воде, не содержащей РНКазы, и количественно определяли с помощью УФ-спектрометрии. мРНК стандартизировали до концентрации 1,0 мкг/мкл и инъецировали 0–1-часовым эмбрионам, полученным из snk 229 /Df(3L)Exel8157 матери, у которых любой продуцируемый белок Snake будет иметь раннее усечение, удаляющее клип и каталитические домены [9]. По соглашению эти материнские мутантные эмбрионы обозначаются как snk -нулевые эмбрионы. После инъекции эмбрионы содержались во влажной камере при 18 °C, как описано [12, 13]. Инъецированные эмбрионы были визуально осмотрены во время ранней гаструляции и оценены как отсутствие спасения, частичное спасение или полное спасение, как описано ([12, 14]; см. Дополнительный рис. 1). Точность этой оценки была проверена путем подготовки кутикулы и окрашивания Twist [15], хотя из-за значительных потерь инъецированных эмбрионов во время этих процедур обычно оценивали только фенотип гаструляции.
Экстрагированную фенолом-хлороформом, осажденную изопропанолом мРНК растворяли в воде, не содержащей РНКазы, и количественно определяли с помощью УФ-спектрометрии. мРНК стандартизировали до концентрации 1,0 мкг/мкл и инъецировали 0–1-часовым эмбрионам, полученным из snk 229 /Df(3L)Exel8157 матери, у которых любой продуцируемый белок Snake будет иметь раннее усечение, удаляющее клип и каталитические домены [9]. По соглашению эти материнские мутантные эмбрионы обозначаются как snk -нулевые эмбрионы. После инъекции эмбрионы содержались во влажной камере при 18 °C, как описано [12, 13]. Инъецированные эмбрионы были визуально осмотрены во время ранней гаструляции и оценены как отсутствие спасения, частичное спасение или полное спасение, как описано ([12, 14]; см. Дополнительный рис. 1). Точность этой оценки была проверена путем подготовки кутикулы и окрашивания Twist [15], хотя из-за значительных потерь инъецированных эмбрионов во время этих процедур обычно оценивали только фенотип гаструляции. Положение инъекции в дорсальную, вентральную или центральную локализованную цитоплазму не влияло на ответ спасения, как описано ранее [9].].
Положение инъекции в дорсальную, вентральную или центральную локализованную цитоплазму не влияло на ответ спасения, как описано ранее [9].].
Функциональный анализ путем совместной трансфекции в клетки Drosophila S2
Дикого типа и четыре мутантных кДНК змеи (R95A, loop1, loop3L, loop3R) вырезали из вектора pBS-SK – с помощью KpnI и NotI и субклонировали в вектор pMT-V5/HisA (Invitrogen) для экспрессии немеченого Snake в культуре клеток Drosophila S2. Ранее были описаны конструкции, кодирующие функциональные HA-меченые GD и FLAG-меченные зимогены Easter [11]. Все эти конструкции экспрессировались под контролем промотора металлотионеина, индуцируемого медью. Культурный 9Клетки 0013 Drosophila S2 (Invitrogen) временно трансфицировали с использованием липофектина (Life Technologies, Grand Island, NY). В параллельные образцы вводили по 1,5 мкг каждой плазмиды (всего до 4,5 мкг), используя пустой вектор для выравнивания общего количества ДНК. После трансфекции (20 ч) клетки промывали бессывороточной средой и экспрессию плазмиды индуцировали 0,7 мМ CuSO 4 . Сбор клеток и сред проводили через 18–20 часов после индукции. Осажденные клетки ресуспендировали в среде путем осторожного пипетирования и аликвоту по 100 мкл каждой клеточной суспензии центрифугировали для разделения клеток и среды. Образцы среды смешивали с равным объемом (100 мкл) буфера для образцов Лэммли, содержащего 100 мМ ДТТ, а образцы клеток ресуспендировали в 200 мкл восстанавливающего буфера для образцов. Эти образцы кипятили в течение 5 минут перед загрузкой в гели.
После трансфекции (20 ч) клетки промывали бессывороточной средой и экспрессию плазмиды индуцировали 0,7 мМ CuSO 4 . Сбор клеток и сред проводили через 18–20 часов после индукции. Осажденные клетки ресуспендировали в среде путем осторожного пипетирования и аликвоту по 100 мкл каждой клеточной суспензии центрифугировали для разделения клеток и среды. Образцы среды смешивали с равным объемом (100 мкл) буфера для образцов Лэммли, содержащего 100 мМ ДТТ, а образцы клеток ресуспендировали в 200 мкл восстанавливающего буфера для образцов. Эти образцы кипятили в течение 5 минут перед загрузкой в гели.
Поликлональные антитела и Вестерн-блоттинг
Анти-SNKCAT были созданы против слитого белка trpE, содержащего 64 аминокислоты (аминокислоты 237–300) в каталитическом домене Snake [9, 16], путем иммунизации кроликов (Pocono Rabbit Farm и Лаборатория). Антитело очищали с помощью положительной селекции против SNKCAT-trpE и отрицательной селекции против trpE, как описано ранее [17, 18], и использовали в рабочей концентрации 1:500 для Вестерн-блоттинга. Ранее были описаны анти-GD-антитела и анти-Easter-антитела, предоставленные Дональдом Морисато [11, 19].] и использовали в концентрациях 1:1500 и 1:2000 соответственно.
Ранее были описаны анти-GD-антитела и анти-Easter-антитела, предоставленные Дональдом Морисато [11, 19].] и использовали в концентрациях 1:1500 и 1:2000 соответственно.
Вестерн-блоты белков, разделенных на 13% гелях SDS-PAGE, исследовали с помощью этих кроличьих поликлональных антител [20]. Хемилюминесцентный субстрат с максимальной чувствительностью SuperSignal West Femto (Pierce) использовался для всех детектирований анти-SNKCAT и для детектирования анти-Easter на рисунке 5, тогда как SuperSignal West Pico (Pierce) или Amersham ECL (GE Healthcare) использовались для остальных экспериментов. . Образцы среды показаны для блотов GD (хотя образцы клеток также анализировались), в то время как образцы клеток показаны для блотов Snake и Easter из-за дефекта секреции мутантных форм Snake и потому, что продукт расщепления Easter является только клеточно-ассоциированным. 11].
Спасательная активность In vivo мутантных форм Snake дикого типа и клип-домена одна тройная мутация мРНК для спасения
snk нулевых эмбрионов. Положения мутантных остатков показаны на двумерной диаграмме домена Snake clip (), где предполагается, что остатки петли 3, расположенные между цистеинами 3 и 4, образуют ободок центральной гидрофобной щели, а остатки петли 1, лежащие между цистеины 1 и 2 могут образовывать заряженный выступ, выступающий перпендикулярно центральной щели [4, 5]. Спасение оценивалось как полное, частичное или отсутствие спасения, что соответствовало паттернам гаструляции дикого типа, частично дорсальной и полностью дорсальной гаструляции соответственно (; см. Методы).
Положения мутантных остатков показаны на двумерной диаграмме домена Snake clip (), где предполагается, что остатки петли 3, расположенные между цистеинами 3 и 4, образуют ободок центральной гидрофобной щели, а остатки петли 1, лежащие между цистеины 1 и 2 могут образовывать заряженный выступ, выступающий перпендикулярно центральной щели [4, 5]. Спасение оценивалось как полное, частичное или отсутствие спасения, что соответствовало паттернам гаструляции дикого типа, частично дорсальной и полностью дорсальной гаструляции соответственно (; см. Методы).Открыть в отдельном окне
Схема домена Snake clip, показывающая мутировавшие остатки. Показана последовательность клип-домена от F92 до P139 (линейное изображение см. в тексте методов) с характерными дисульфидными связями (C 1 -C 5 ; C 2 -C 4 ; C 3 -C). 6 ) и указанное расположение петель 1 и 3. Заряженные остатки, которые индивидуально и в группах мутировали в аланин, обозначены черными кружками. Одна звездочка идентифицирует R95, мутация привела к небольшому дефекту в активности по спасению эмбрионов, в то время как двойные звездочки обозначают остатки R94 и D98, которые оба были мутированы в мутанте loop1, что привело к серьезному дефекту в активности по спасению эмбрионов.
Одна звездочка идентифицирует R95, мутация привела к небольшому дефекту в активности по спасению эмбрионов, в то время как двойные звездочки обозначают остатки R94 и D98, которые оба были мутированы в мутанте loop1, что привело к серьезному дефекту в активности по спасению эмбрионов.
Таблица 1
Фенотипическое спасение с помощью мутантов Snake
| Конструкция | Номер | Полный Rescue (%) | Partquial Rescue (%) | ).|
|---|---|---|---|---|
| WT | 272 | 165(61) | 61(22) | 46(17) |
| SnkR94A | 64 | 45(70) | 11(17) | 8( 13) |
| SnkR95A | 430 | 146(34) | 129(30) | 155(36) |
| SnkD98A | 62 | 33(53) | 11(18) | 18(29) |
| СнкР100А | 97 | 67(69) | 19(20) | 11(11) |
| SnkR115A | 94 | 56(60) | 24(25) | 14(15) |
| SnkE116A | 64 | 48(75) | 9(14) | 7(11) |
| SnkR118A | 74 | 38(51) | 10(14) | 26(35) |
| SnkR123A | 36 | 21(58) | 10(28) | 5(14) |
| SnkD125A | 47 | 25(53) | 14(30) | 8(17) |
| SnkR130A | 44 | 30(68) | 6(14) | 8(18 ) |
| loop1 | 485 | 20(4) | 50(10) | 415(86) |
| loop3L | 49 | 23(47) | 18(37) | 8 (16) |
| петля3R | 140 | 88(62) | 26(19) | 26(19) |
Открыть в отдельном окне
Основным выводом этого исследования было то, что только один из тринадцати мутантов, loop1 (R94A, D98A), проявлял серьезные нарушения функционируют в тесте на спасение, при этом только 4% эмбрионов полностью спасаются, а 86% остаются полностью дорсальными. Напротив, большинство однократных изменений заряда в аланин спасли нулевые эмбрионы snk- с эффективностью, приблизительно равной эффективности дикого типа. Среди одиночных мутантов были отмечены два скромных недостатка в спасательной деятельности. Во-первых, R9.Мутант 5A показал пониженную способность обеспечивать полное спасение (34% по сравнению с 61% для дикого типа). Во-вторых, мутант R95A, а также мутанты D98A и R118A показали увеличение процента не спасенных эмбрионов с полностью дорсальным фенотипом (36%, 29% и 35% соответственно по сравнению с 17% для мРНК змеи дикого типа ). Эти результаты согласуются с задержкой или снижением способности Снейка достичь необходимого порога активности в течение ограниченного периода времени, когда может произойти спасение. Тройной мутант loop3L, который сочетает в себе R118A с R115A и E116A, работал лучше, чем R118A, со сдвигом в сторону частичного восстановления (37% по сравнению с 14% для R118A) и аналогичного полного восстановления, что позволяет предположить, что изменения в плотности заряда могут влиять на домен клипа.
Напротив, большинство однократных изменений заряда в аланин спасли нулевые эмбрионы snk- с эффективностью, приблизительно равной эффективности дикого типа. Среди одиночных мутантов были отмечены два скромных недостатка в спасательной деятельности. Во-первых, R9.Мутант 5A показал пониженную способность обеспечивать полное спасение (34% по сравнению с 61% для дикого типа). Во-вторых, мутант R95A, а также мутанты D98A и R118A показали увеличение процента не спасенных эмбрионов с полностью дорсальным фенотипом (36%, 29% и 35% соответственно по сравнению с 17% для мРНК змеи дикого типа ). Эти результаты согласуются с задержкой или снижением способности Снейка достичь необходимого порога активности в течение ограниченного периода времени, когда может произойти спасение. Тройной мутант loop3L, который сочетает в себе R118A с R115A и E116A, работал лучше, чем R118A, со сдвигом в сторону частичного восстановления (37% по сравнению с 14% для R118A) и аналогичного полного восстановления, что позволяет предположить, что изменения в плотности заряда могут влиять на домен клипа. действуют тонкими способами. Скромные недостатки, замеченные для R95A и D98A, вместе с тяжелым дефицитом функции двойного мутанта loop1 (R94A, D98A), предполагают, что предсказанная выступающая область петли 1 играет важную роль в функции Snake in vivo (14).
действуют тонкими способами. Скромные недостатки, замеченные для R95A и D98A, вместе с тяжелым дефицитом функции двойного мутанта loop1 (R94A, D98A), предполагают, что предсказанная выступающая область петли 1 играет важную роль в функции Snake in vivo (14).
Помимо изучения влияния мутаций клипового домена на активность полноразмерного белка Snake, мы стремились определить, будут ли продомены, содержащие эти мутации, иметь сниженную способность к дорсализации эмбрионов дикого типа, как сообщалось для продомена дикого типа [ 9]. Эти авторы предположили, что продомены Snake и Easter могут конкурировать с полноразмерными белками за взаимодействия с др. протеазами или кофакторами в этом пути. Однако нам не удалось воспроизвести опубликованные данные о доминантно-негативной активности продомена Snake дикого типа, используя либо собственные 5′- и 3′-UTR Snake, либо заменив Xenopus β-globin 5’UTR и easter 3’UTR (в попытке повысить эффективность трансляции мРНК) и с использованием последовательности продомена с цистеином или без него 179 мутировали в серин (поскольку неясно, какой из них использовался в оригинальном исследовании, и свободный цистеин 179 мог привести к аномальному дисульфидному связыванию в отсутствие каталитического домена [23, 24]) (данные не показаны). Поэтому мы не проводили эти эксперименты с конструкциями мутантных продоменов.
Поэтому мы не проводили эти эксперименты с конструкциями мутантных продоменов.
Взаимодействие мутантных белков Snake с другими компонентами каскада в культивируемых клетках дрозофилы S2
Было показано, что дорсовентральные протеазы GD, Snake и Easter функционируют в последовательном каскаде активации зимогена при совместной экспрессии либо в клетках Drosophila S2, либо в бакуловирус-инфицированных клетках чешуекрылых [10, 11]. Эти анализы на основе клеточных культур вряд ли точно имитируют взаимодействия протеаз in vivo, например, процессинг, по-видимому, происходит только в секреторном пути коэкспрессирующих клеток, а не в кондиционированных средах; однако они демонстрируют основную каталитическую функцию протеаз, а также ряд потенциальных расщеплений по принципу обратной связи между протеазами. Значение in vivo этих расщеплений по принципу обратной связи, наблюдаемых в клеточной культуре, неизвестно.
Взаимодействия протеаз, обнаруженные в системе коэкспрессии клеток S2 [11], кратко описаны здесь (). Совместная экспрессия зимогенов GD и Snake приводит к перекрестному процессингу обоих ферментов. Либо низкая активность зимогена GD, либо небольшое количество обработанного активного GD, генерируемого неспецифически в секреторном пути, по-видимому, способны расщеплять Snake в месте расщепления его зимогена (реакция 1), и, в свою очередь, активированный Snake может процессировать зимоген GD. (реакция 2), создавая петлю положительной обратной связи. Активированный Snake может также процессировать другие молекулы Snake в некартированном сайте внутри продомена (реакция 3; [25]), так что большой C-концевой фрагмент Snake является преобладающим продуктом процессинга в котрансфекциях GD/Snake (, полосы 2 и 4). Если также присутствует пасхальный зимоген (тройная трансфекция), он активируется Снейком (реакция 4) и, в свою очередь, может обрабатывать GD в той же области, что и Снейк (реакция 5), что может привести к дополнительной положительной обратной связи (или отрицательный, как предложено [10]).
Совместная экспрессия зимогенов GD и Snake приводит к перекрестному процессингу обоих ферментов. Либо низкая активность зимогена GD, либо небольшое количество обработанного активного GD, генерируемого неспецифически в секреторном пути, по-видимому, способны расщеплять Snake в месте расщепления его зимогена (реакция 1), и, в свою очередь, активированный Snake может процессировать зимоген GD. (реакция 2), создавая петлю положительной обратной связи. Активированный Snake может также процессировать другие молекулы Snake в некартированном сайте внутри продомена (реакция 3; [25]), так что большой C-концевой фрагмент Snake является преобладающим продуктом процессинга в котрансфекциях GD/Snake (, полосы 2 и 4). Если также присутствует пасхальный зимоген (тройная трансфекция), он активируется Снейком (реакция 4) и, в свою очередь, может обрабатывать GD в той же области, что и Снейк (реакция 5), что может привести к дополнительной положительной обратной связи (или отрицательный, как предложено [10]). Активированный Easter также надежно расщепляет зимоген Snake в продомене в некартированном сайте C-конца к Snake-зависимому сайту процессинга (реакция 6; [11] и , дорожки 1 и 5). Сходные события были описаны в инфицированных бакуловирусом клетках чешуекрылых, за исключением того, что активация GD при коэкспрессии со Snake оказалась автокаталитической, требующей присутствия Snake, но не требующей каталитической активности Snake [10]. Причина этого различия между двумя системами коэкспрессии не была исследована, однако возможно, что отдельная протеаза чешуекрылых способна активировать зимоген GD, когда он находится в комплексе со Snake.
Активированный Easter также надежно расщепляет зимоген Snake в продомене в некартированном сайте C-конца к Snake-зависимому сайту процессинга (реакция 6; [11] и , дорожки 1 и 5). Сходные события были описаны в инфицированных бакуловирусом клетках чешуекрылых, за исключением того, что активация GD при коэкспрессии со Snake оказалась автокаталитической, требующей присутствия Snake, но не требующей каталитической активности Snake [10]. Причина этого различия между двумя системами коэкспрессии не была исследована, однако возможно, что отдельная протеаза чешуекрылых способна активировать зимоген GD, когда он находится в комплексе со Snake.
Открыть в отдельном окне
Обзор расщеплений, обнаруженных среди дорсовентральных протеаз в системе анализа коэкспрессии клеток S2. (А) Схематическая диаграмма расщеплений, происходящих при совместной экспрессии GD и Snake (реакции 1–3) или при совместной экспрессии GD, Snake и Easter (реакции 1–6) [11]. Описание этих реакций см. в тексте результатов. (B) Обработка каталитического серин-мутированного субстрата Snake (SNKS-A) предварительно активированными (ΔN) формами Easter, Snake и GD приводит к различным C-концевым продуктам (дорожки 1–3). Зависимый от GD процессинг был картирован с сайтом расщепления зимогена (продукт отмечен стрелкой), в то время как сайты процессинга, зависящие от Easter и Snake, находятся в некартированных позициях в продомене Snake. При двойной трансфекции GD и функционального Snake преобладает Snake-зависимое расщепление (дорожка 4), тогда как при тройной трансфекции этих протеаз плюс Easter преобладает Easter-зависимый продукт расщепления (дорожка 5). Дорожки 1, 3 и 5 были представлены ранее на рисунке 1B в [11] и включены сюда для сравнения с дорожками 2 и 4, документирующими зависимое от змеи расщепление змеи из того же геля. В отличие от — в этой статье, в которой используется немеченый Snake, распознаваемый антителом с каталитическим доменом Snake, субстрат SNKS-A включает C-концевую 5-myc-6-His-метку ~ 10 кДа и был обнаружен с использованием антитела против myc.
(B) Обработка каталитического серин-мутированного субстрата Snake (SNKS-A) предварительно активированными (ΔN) формами Easter, Snake и GD приводит к различным C-концевым продуктам (дорожки 1–3). Зависимый от GD процессинг был картирован с сайтом расщепления зимогена (продукт отмечен стрелкой), в то время как сайты процессинга, зависящие от Easter и Snake, находятся в некартированных позициях в продомене Snake. При двойной трансфекции GD и функционального Snake преобладает Snake-зависимое расщепление (дорожка 4), тогда как при тройной трансфекции этих протеаз плюс Easter преобладает Easter-зависимый продукт расщепления (дорожка 5). Дорожки 1, 3 и 5 были представлены ранее на рисунке 1B в [11] и включены сюда для сравнения с дорожками 2 и 4, документирующими зависимое от змеи расщепление змеи из того же геля. В отличие от — в этой статье, в которой используется немеченый Snake, распознаваемый антителом с каталитическим доменом Snake, субстрат SNKS-A включает C-концевую 5-myc-6-His-метку ~ 10 кДа и был обнаружен с использованием антитела против myc. . Приблизительные молекулярные массы полос, связанных со змеей, указаны слева, а маркеры молекулярной массы указаны справа в этом и будущих пятнах.
. Приблизительные молекулярные массы полос, связанных со змеей, указаны слева, а маркеры молекулярной массы указаны справа в этом и будущих пятнах.
Для дальнейшего изучения влияния мутаций клип-домена Snake на функцию Snake мы протестировали четыре из этих мутантов — R95A, loop1, loop3L и loop3R — в системе экспрессии клеток S2. Эти четыре мутанта были выбраны потому, что мутанты с одинарной петлей R95A и двойной петлей 1 демонстрировали умеренные или серьезные дефекты, соответственно, в спасательной активности путем инъекции эмбриона, поэтому могли демонстрировать дефекты в специфических взаимодействиях в анализах коэкспрессии, в то время как мутанты с тройной петлей 3L и двойной петлей 3R этого не делали. не обнаруживали дефектов в тесте с инъекцией эмбриона, и ожидалось, что они будут хорошо работать в исследованиях коэкспрессии. При экспрессии по отдельности все четыре мутанта демонстрировали сильный дефект секреции по сравнению с Snake дикого типа, но присутствовали на нормальных уровнях без продуктов деградации (26), что указывает на дефект переноса мутантных белков Snake через секреторный путь [26]. Поскольку процессинг коэкспрессируемых зимогенов происходит внутри секреторного пути, а не в среде [11], мы приступили к тестированию мутантов, предполагая, что соответствующие взаимодействия все еще могут происходить внутриклеточно. Для простоты петля1 и R95A результаты всех котрансфекций представлены перед результатами loop3L и loop3R.
Поскольку процессинг коэкспрессируемых зимогенов происходит внутри секреторного пути, а не в среде [11], мы приступили к тестированию мутантов, предполагая, что соответствующие взаимодействия все еще могут происходить внутриклеточно. Для простоты петля1 и R95A результаты всех котрансфекций представлены перед результатами loop3L и loop3R.
Открыть в отдельном окне
Дефект секреции мутантов Snake и дефекты процессинга, наблюдаемые при котрансфекции зимогеном GD дикого типа. (A) Все протестированные мутанты (loop3R, loop1, R95A и loop3L) имеют дефект секреции по сравнению со Snake дикого типа (WT) в клетках S2. М и С обозначают среду и клетки соответственно. (B) Котрансфекция GD пустым вектором pMT или змеей дикого типа или мутантной. IB, антитело для иммуноблоттинга. Верхняя панель, петля1 и R9Мутанты змеи 5A демонстрируют переменный и умеренно сниженный процессинг зимогена GD (GDz) до продуктов его С-концевого расщепления (GDc, дублет 46–50 кДа) по сравнению со змеей дикого типа, в то время как loop3R и loop3L демонстрируют большее снижение процессинга GD. Нижняя панель, мутанты R95A и loop1 демонстрируют нормальный Snake-зависимый процессинг змеиного зимогена (SNKz) до С-концевого продукта массой 43 кДа (SNKcs, где s указывает Snake-зависимость этой формы), в то время как loop3R и loop3L демонстрируют умеренное и сильное снижение обработки соответственно.
Нижняя панель, мутанты R95A и loop1 демонстрируют нормальный Snake-зависимый процессинг змеиного зимогена (SNKz) до С-концевого продукта массой 43 кДа (SNKcs, где s указывает Snake-зависимость этой формы), в то время как loop3R и loop3L демонстрируют умеренное и сильное снижение обработки соответственно.
Удивительно, но по сравнению с анализом по спасению эмбрионов мутантная петля loop1 довольно хорошо проявляла себя в отношении расщеплений, обнаруживаемых в клеточной системе S2. При двойной трансфекции зимогенов GD и Snake () наблюдался устойчивый процессинг в продомене Snake, который, как известно, осуществляется Snake, а не GD (реакция 3; , дорожки 2 и 3) и зависит от предшествующей активации Snake GD (не показано). при однократной трансфекции змеи [11]). Наблюдалось вариабельное снижение Snake-зависимого процессинга GD при котрансфекциях с петлей 1 или R9.5A мутант Snake по сравнению с диким типом (реакция 2; и данные не показаны), при этом мутант loop1 более затронут, чем R95A. Однако при тройной трансфекции как зимогенами GD, так и зимогенами Easter не было обнаружено различий между Snake дикого типа и змеей с мутантной петлей R95A в отношении расщепления любой из протеаз (1). Эти результаты вместе предполагают, что мутации петли 1 не показывают функционально значимых нарушений каталитической активности Snake или взаимодействия с GD и Easter при анализе в клеточной системе S2, и что способность Easter обрабатывать GD может маскировать скромный дефект, наблюдаемый для процессинга петли 1 GD. при двойной трансфекции.
Однако при тройной трансфекции как зимогенами GD, так и зимогенами Easter не было обнаружено различий между Snake дикого типа и змеей с мутантной петлей R95A в отношении расщепления любой из протеаз (1). Эти результаты вместе предполагают, что мутации петли 1 не показывают функционально значимых нарушений каталитической активности Snake или взаимодействия с GD и Easter при анализе в клеточной системе S2, и что способность Easter обрабатывать GD может маскировать скромный дефект, наблюдаемый для процессинга петли 1 GD. при двойной трансфекции.
Открыть в отдельном окне
Дефекты процессинга в значительной степени маскируются при тройной трансфекции GD, Easter и мутантными зимогенами Snake. Верхняя панель, ГД. Средняя панель, Змея, с указанием продукта процессинга SNKze (32 кДа), зависящего от Пасхи. Нижняя панель, Пасха, показывает небольшое снижение процессинга мутантом loop3L и умеренное снижение процессинга мутантом loop3R.
Ранее предполагалось, что clip-домен Snake блокирует каталитический сайт протеазного домена, предотвращая Snake расщепление Easter, за исключением случаев, когда Snake присутствует в вентральном активирующем комплексе [25]. Если бы мутант loop1 утратил эту ингибирующую роль, протеаза Snake могла бы слабо, но без разбора активировать Easter во внеклеточном пространстве эмбриона, приводя к слабой латерализации или вентрализации эмбриона. В качестве одного из тестов этой возможности in vitro мы выполнили двойную трансфекцию Snake и Easter (2), чтобы увидеть, можно ли обнаружить повышенный процессинг Easter в мутанте loop1 по сравнению с диким типом. В то время как в предыдущих исследованиях не было обнаружено никакого процессинга Easter при таких двойных трансфекциях [11], с помощью более чувствительного хемилюминесцентного реагента мы обнаружили небольшое количество процессинга Easter, которое зависело от коэкспрессии Snake. Петля1 и R95A мутанты демонстрировали аналогичный или меньший процессинг по сравнению со Snake дикого типа, поэтому мы не смогли найти подтверждение роли этих остатков клип-домена в блокировании каталитического сайта Snake.
Если бы мутант loop1 утратил эту ингибирующую роль, протеаза Snake могла бы слабо, но без разбора активировать Easter во внеклеточном пространстве эмбриона, приводя к слабой латерализации или вентрализации эмбриона. В качестве одного из тестов этой возможности in vitro мы выполнили двойную трансфекцию Snake и Easter (2), чтобы увидеть, можно ли обнаружить повышенный процессинг Easter в мутанте loop1 по сравнению с диким типом. В то время как в предыдущих исследованиях не было обнаружено никакого процессинга Easter при таких двойных трансфекциях [11], с помощью более чувствительного хемилюминесцентного реагента мы обнаружили небольшое количество процессинга Easter, которое зависело от коэкспрессии Snake. Петля1 и R95A мутанты демонстрировали аналогичный или меньший процессинг по сравнению со Snake дикого типа, поэтому мы не смогли найти подтверждение роли этих остатков клип-домена в блокировании каталитического сайта Snake.
Открыть в отдельном окне
Повышенного процессинга Пасхи при двойной трансфекции мутантными зимогенами Змеи нет. Двойные трансфекции Snake/Easter сравнивают с тройными трансфекциями GD/Snake/Easter, показывающими полный процессинг зимогена Easter, и с двойной трансфекцией Easter/pMT, показывающей отсутствие процессинга. Верхняя панель, при этих двойных трансфекциях двух зимогенов происходит минимальная обработка Snake. Значение слегка отличающихся продуктов переработки у некоторых мутантов по сравнению со змеей дикого типа неизвестно. Нижняя панель: небольшое количество пасхи преобразуется в активированную форму (EAc) зависимым от змеи способом с помощью R9 дикого типа.5A и loop1, но не мутантами loop3R или loop3L.
Двойные трансфекции Snake/Easter сравнивают с тройными трансфекциями GD/Snake/Easter, показывающими полный процессинг зимогена Easter, и с двойной трансфекцией Easter/pMT, показывающей отсутствие процессинга. Верхняя панель, при этих двойных трансфекциях двух зимогенов происходит минимальная обработка Snake. Значение слегка отличающихся продуктов переработки у некоторых мутантов по сравнению со змеей дикого типа неизвестно. Нижняя панель: небольшое количество пасхи преобразуется в активированную форму (EAc) зависимым от змеи способом с помощью R9 дикого типа.5A и loop1, но не мутантами loop3R или loop3L.
В отличие от относительно надежной работы мутантов loop1 и R95A в клеточной системе S2, мутанты loop3L и loop3R показали обширные дефекты, которые были неожиданными, учитывая их способность спасать snk -нулевые эмбрионы. При двойной трансфекции Snake и GD () наблюдалось умеренное снижение процессинга GD и умеренное или сильное снижение процессинга Snake, в то время как при двойной трансфекции Snake и Easter () не было заметного процессинга Easter этими мутантами Snake. Однако, как видно для мутанта loop1, когда все три протеазы экспрессировались вместе (), присутствие Easter, по-видимому, компенсировало недостатки мутантов петли 3 с явно полным восстановлением процессинга GD и специфичного для Easter Snake-процессинга, и несколько снижен процессинг Easter мутантами Snake loop3R и loop3L.
Однако, как видно для мутанта loop1, когда все три протеазы экспрессировались вместе (), присутствие Easter, по-видимому, компенсировало недостатки мутантов петли 3 с явно полным восстановлением процессинга GD и специфичного для Easter Snake-процессинга, и несколько снижен процессинг Easter мутантами Snake loop3R и loop3L.
Ни одна замена заряженного на аланин в домене клипа, а только одна из трех множественных замен не привела к резкому снижению способности зимогена змеи спасать змеиных нулевых эмбрионов. Сходные данные о том, что большинство одиночных изменений явно не влияют на функцию, являются общими в исследованиях мутагенеза заряженных в аланин [22, 26], и для любого отдельного остатка может означать либо то, что он не важен для функции Snake, либо множественные взаимодействия, происходящие с партнер по связыванию маскирует эффект удаления одного заряженного остатка, участвующего в этих взаимодействиях. За исключением двойного мутанта loop1, наблюдаемое спасение эмбриона было относительно сильным ввиду дефекта секреции, наблюдаемого у четырех репрезентативных мутантов, экспрессированных в клеточной культуре. Такие комбинации дефекта секреции с относительно нормальной функцией клеточной поверхности или очищенных белков были описаны ранее для заряженных к аланину мутантов [26]. В случае Snake возможно, что (а) секреторная система эмбриона более эффективна в доставке Snake на клеточную поверхность, чем клетки S2, (b) очень небольшое количество мутантного белка Snake во внеклеточном пространстве достаточно для спасения каскада in vivo, и/или (c) кофактор и/или взаимодействия обратной связи внутри каскада in vivo способны компенсировать снижение количества или качества Snake во внеклеточном пространстве.
Такие комбинации дефекта секреции с относительно нормальной функцией клеточной поверхности или очищенных белков были описаны ранее для заряженных к аланину мутантов [26]. В случае Snake возможно, что (а) секреторная система эмбриона более эффективна в доставке Snake на клеточную поверхность, чем клетки S2, (b) очень небольшое количество мутантного белка Snake во внеклеточном пространстве достаточно для спасения каскада in vivo, и/или (c) кофактор и/или взаимодействия обратной связи внутри каскада in vivo способны компенсировать снижение количества или качества Snake во внеклеточном пространстве.
По-видимому, противоречивые результаты были получены в анализе с инъекцией эмбриона по сравнению с анализом коэкспрессии S2, в котором мутантная петля loop1 показала плохие результаты in vivo, но хорошие результаты in vitro, в то время как мутанты loop3L и loop3R показали хорошие результаты in vivo, но плохие результаты in vitro. Имеющиеся данные предполагают три возможности ухудшения контура 1. Первым и наименее интересным является то, что мутант loop1 может иметь особенно серьезный дефект секреции у эмбрионов и, таким образом, быть недоступным для участия в протеазном каскаде внутри перивителлиновой жидкости. Вторая возможность состоит в том, что мутант loop1 имеет нарушенные взаимодействия с GD, на что намекает незначительно сниженный процессинг GD при двойных трансфекциях (14). Уменьшение взаимодействия GD и Snake in vivo может напрямую привести к снижению активации Snake, снижению предполагаемой активации GD по обратной связи [10, 11] или непосредственному влиянию Snake на активацию Easter, если Snake и GD должны оставаться связанными вместе в вентральном комплексе для активации Easter. произойти [25, 27]. Наконец, уже известно, что клеточная система S2, включающая сверхэкспрессию протеаз, обходит in vivo потребность в вентральном кофакторе, действующем при активации Snake и/или активации Easter [11, 19].]. Т.о., возможно, что критический дефект у мутанта loop1 заключается во взаимодействиях с этим вентральным кофактором, который может концентрировать зимогены протеазы на поверхности.
Первым и наименее интересным является то, что мутант loop1 может иметь особенно серьезный дефект секреции у эмбрионов и, таким образом, быть недоступным для участия в протеазном каскаде внутри перивителлиновой жидкости. Вторая возможность состоит в том, что мутант loop1 имеет нарушенные взаимодействия с GD, на что намекает незначительно сниженный процессинг GD при двойных трансфекциях (14). Уменьшение взаимодействия GD и Snake in vivo может напрямую привести к снижению активации Snake, снижению предполагаемой активации GD по обратной связи [10, 11] или непосредственному влиянию Snake на активацию Easter, если Snake и GD должны оставаться связанными вместе в вентральном комплексе для активации Easter. произойти [25, 27]. Наконец, уже известно, что клеточная система S2, включающая сверхэкспрессию протеаз, обходит in vivo потребность в вентральном кофакторе, действующем при активации Snake и/или активации Easter [11, 19].]. Т.о., возможно, что критический дефект у мутанта loop1 заключается во взаимодействиях с этим вентральным кофактором, который может концентрировать зимогены протеазы на поверхности. Такая роль клип-доменов ранее была предложена для связывания с поверхностями патогенов, таким образом, концентрируя компоненты каскада активации профенолоксидазы для обеспечения локализованной амплификации сигнала [4, 5].
Такая роль клип-доменов ранее была предложена для связывания с поверхностями патогенов, таким образом, концентрируя компоненты каскада активации профенолоксидазы для обеспечения локализованной амплификации сигнала [4, 5].
Зимогены Snake, содержащие множественные мутации заряженного аланина в петле 3, которые, вероятно, структурно соответствуют открытому краю центральной щели в домене clip, относятся к числу мутантов, выполняющих функцию дикого типа in vivo, однако они проявляют тяжелые нарушение Snake-зависимых реакций процессинга при двойной трансфекции либо GD, либо Easter (, ). Это нарушение предполагает, что каталитический домен Снейка имеет очень низкую активность в этих экспериментальных условиях. Это может быть результатом нарушения расщепления зимогена Snake или снижения способности принимать открытую и активную конформацию каталитического домена Snake после расщепления зимогена, например, если предложенная блокировка каталитического сайта клип-доменом Snake [25] не исчезает при мутации остатков петли 3. Интересно, что эти дефекты маскируются в тройных трансфекциях, содержащих помимо GD и Snake (20) Истер. Расщепление Easter при тройных трансфекциях строго зависит от каталитической активности Snake [11], что свидетельствует о том, что каталитический домен Snake уложен правильно и может функционировать в мутантах loop3L и loop3R. Продукции небольшого количества активного Easter может быть достаточно, чтобы сделать возможным сильное зависимое от Easter расщепление GD и Snake, наблюдаемое при тройной трансфекции, особенно если последнее расщепление внутри продомена Snake приводит к снятию ингибирования каталитического домена Snake с помощью мутантные домены клипа, чтобы обеспечить более нормальный уровень активации Easter. Альтернативная возможность, предложенная этими результатами, заключается в том, что GD, Snake и Easter лучше всего функционируют во взаимно активирующемся комплексе, и что множественные межмолекулярные взаимодействия между этими протеазами преодолевают дефекты петли 3 как в тройных трансфекциях клеток S2, так и в анализах с инъекцией эмбриона.
Интересно, что эти дефекты маскируются в тройных трансфекциях, содержащих помимо GD и Snake (20) Истер. Расщепление Easter при тройных трансфекциях строго зависит от каталитической активности Snake [11], что свидетельствует о том, что каталитический домен Snake уложен правильно и может функционировать в мутантах loop3L и loop3R. Продукции небольшого количества активного Easter может быть достаточно, чтобы сделать возможным сильное зависимое от Easter расщепление GD и Snake, наблюдаемое при тройной трансфекции, особенно если последнее расщепление внутри продомена Snake приводит к снятию ингибирования каталитического домена Snake с помощью мутантные домены клипа, чтобы обеспечить более нормальный уровень активации Easter. Альтернативная возможность, предложенная этими результатами, заключается в том, что GD, Snake и Easter лучше всего функционируют во взаимно активирующемся комплексе, и что множественные межмолекулярные взаимодействия между этими протеазами преодолевают дефекты петли 3 как в тройных трансфекциях клеток S2, так и в анализах с инъекцией эмбриона.
Наши результаты демонстрируют важность заряженных остатков петли 1 (R94, R95, D98) в clip-домене для функции Snake in vivo и позволяют предположить, что остатки петли 3 могут регулировать каталитическую функцию Snake в некоторых экспериментальных условиях. Противоречивые результаты, полученные в анализе инъекции эмбриона и анализе коэкспрессии S2, подчеркивают важность использования нескольких анализов и необходимость осторожности при интерпретации данных любого отдельного анализа функции протеазы. Чтобы определить, функционирует ли домен клипсы Snake как модуль белок-белкового взаимодействия для связывания либо с GD, либо с вентральным кофактором, или если он регулирует каталитическую функцию Snake, потребуется разработка как анализов in vitro с использованием очищенных протеаз, так и более совершенных инструментов для изучения протеаз. Активация и локализация in vivo. Вопросы, доступные с использованием текущих анализов, включают картирование сайтов в Snake, которые обрабатываются Snake и Easter, и определение их значимости для протеазного каскада, а также выполнение мутагенеза заряженного аланина домена клипа Easter, чтобы увидеть, есть ли мутации в петле 1 и петле 3. демонстрируют поведение, аналогичное тому, которое наблюдалось у Змеи.
демонстрируют поведение, аналогичное тому, которое наблюдалось у Змеи.
01
Нажмите здесь для просмотра. (35K, документ)
02
Нажмите здесь для просмотра. (1.0M, tif)
Мы благодарим Maggie Elalayli и Jacklyn Hall за помощь в создании антитела против SNKCAT; Роберт ДеЛотто, Дэвид Стейн, Дональд Морисато, Зигфрид Рот, Блумингтонский фондовый центр и Exelixis за реагенты и запасы мух; и анонимных рецензентов за их предложения. Эта работа была поддержана NIH RO1 GM067738 до E.K.L.
- PPAF-I or –II
- prophenoloxidase-activating factor I or II
- PAP-2
- prophenoloxidase-activating proteinase-2
- GD
- Gastrulation Defective
- SNK
- Snake
- EA
- Easter
Отказ от ответственности издателя: Это PDF-файл неотредактированной рукописи, принятой к публикации. В качестве услуги нашим клиентам мы предоставляем эту раннюю версию рукописи. Рукопись будет подвергнута редактированию, набору текста и рецензированию полученного доказательства, прежде чем она будет опубликована в ее окончательной цитируемой форме. Обратите внимание, что в процессе производства могут быть обнаружены ошибки, которые могут повлиять на содержание, и все правовые оговорки, применимые к журналу, относятся к нему.
Рукопись будет подвергнута редактированию, набору текста и рецензированию полученного доказательства, прежде чем она будет опубликована в ее окончательной цитируемой форме. Обратите внимание, что в процессе производства могут быть обнаружены ошибки, которые могут повлиять на содержание, и все правовые оговорки, применимые к журналу, относятся к нему.
1. Цзян Х., Каност М.Р. Насекомое Биохим Мол Биол. 2000;30:95–105. [PubMed] [Google Scholar]
2. Росс Дж., Цзян Х., Каност М.Р., Ван Ю. Джин. 2003; 304: 117–131. [PubMed] [Google Scholar]
3. Muta T, Hashimoto R, Miyata T, Nishimura H, Toh Y, Iwanaga S. J Biol Chem. 1990; 265:22426–22433. [PubMed] [Google Scholar]
4. Хуан Р., Лу З., Дей Х., Вандер Вельде Д., Пракаш О., Цзян Х. Биохимия. 2007;46:11431–11439. [PubMed] [Google Scholar]
5. Piao S, Song YL, Kim JH, Park SY, Park JW, Lee BL, Oh BH, Ha NC. EMBO J. 2005; 24:4404–4414. [Бесплатная статья PMC] [PubMed] [Google Scholar]
6. Moussian B, Roth S. Curr Biol. 2005; 15: Р887–Р899. [PubMed] [Google Scholar]
Curr Biol. 2005; 15: Р887–Р899. [PubMed] [Google Scholar]
7. Часан Р., Андерсон К.В. Клетка. 1989; 56: 391–400. [PubMed] [Google Scholar]
8. ДеЛотто Р., Шпирер П. Природа. 1986; 323: 688–692. [PubMed] [Google Scholar]
9. Smith C, Giordano H, DeLotto R. Генетика. 1994; 136:1355–1365. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Dissing M, Giordano H, DeLotto R. EMBO J. 2001;20:2387–2393. [Бесплатная статья PMC] [PubMed] [Google Scholar]
11. LeMosy EK, Tan YQ, Hashimoto C. Proc Natl Acad Sci USA. 2001; 98: 5055–5060. [Статья бесплатно PMC] [PubMed] [Google Scholar]
12. Rose T, LeMosy EK, Cantwell AM, Banerjee-Roy D, Skeath JB, Di Cera E. J Biol Chem. 2003; 278:11320–11330. [PubMed] [Google Scholar]
13. Misra S, Hecht P, Maeda R, Anderson KV. Разработка. 1998; 125:1261–1267. [PubMed] [Google Scholar]
14. Андерсон К.В., Юргенс Г., Нусляйн-Фольхард К. Селл. 1985; 42: 779–789. [PubMed] [Академия Google]
15. Stein D, Roth S, Vogelsang E, Nüsslein-Volhard C. Cell. 1991; 65: 725–735. [PubMed] [Google Scholar]
Stein D, Roth S, Vogelsang E, Nüsslein-Volhard C. Cell. 1991; 65: 725–735. [PubMed] [Google Scholar]
16. Koerner TJ, Hill JE, Myers AM, Tzagaloff A. Meth Enzymol. 1990; 194: 477–490. [PubMed] [Google Scholar]
17. Хашимото С., Хадсон К.Л., Андерсон К.В. Разработка. 1991; 111:1021–1028. [PubMed] [Google Scholar]
18. LeMosy EK, Kemler D, Hashimoto C. Development. 1998; 125:4045–4053. [PubMed] [Google Scholar]
19. LeMosy EK. ФЭБС лат. 2006;580:2269–2272. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Harlow E, Lane D. Использование антител: лабораторное руководство. Лабораторный пресс Колд-Спринг-Харбор; Plainview, NY: 1999. [Google Scholar]
21. Gibbs CS, Zoller MJ. Дж. Биол. Хим. 1991; 266:8923–8931. [PubMed] [Google Scholar]
22. Bass SH, Mulkerrin MG, Wells JA. Proc Natl Acad Sci USA. 1991; 88: 4498–4502. [Бесплатная статья PMC] [PubMed] [Google Scholar]
23. Smith CL, DeLotto R. Nature. 1994; 368: 548–551. [PubMed] [Академия Google]
24. Часан Р., Джин И., Андерсон К.В. Разработка. 1992; 115: 607–616. [PubMed] [Google Scholar]
Часан Р., Джин И., Андерсон К.В. Разработка. 1992; 115: 607–616. [PubMed] [Google Scholar]
25. Smith CL, Giordano H, Schwartz M, DeLotto R. Development. 1995; 121:4127–4135. [PubMed] [Google Scholar]
26. Duchesnes CE, Murphy PM, Williams TJ, Pease JE. Мол Иммунол. 2006;43:1221–1231. [PubMed] [Google Scholar]
27. Пономарев Г., Джордано Х., ДеЛотто Ю., ДеЛотто Р. Генетика. 2001; 159: 635–645. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9Плакат 0000 Bluz Gdz 2020 — Годзилла Кинг КонгДизайн также доступен на
Вместе с этим часто покупают
Узнать большеВам также может понравиться
- Дом
- Футболки
- Годзилла Кинг Конг Футболки
- Плакат Bluz Gdz 2020 — Годзилла Кинг Конг — Футболка
Особенности
Горячая тенденция, на которую вы смотрите, это Плакат Bluz gdz 2020 — Годзилла Кинг-Конг — Футболка Для Мужчины Черный Годзилла Кинг Конг Футболки в Printerval:
Из какого материала сделан этот товар?
Материал может отличаться для разных стилей или продуктов и цветов. Учить больше»
Учить больше»
Какова ваша политика возврата?
Мы хотим, чтобы вы любили свой заказ! Если по какой-либо причине вы этого не сделаете, сообщите нам, и мы все исправим. Учить больше»
Какова ваша политика возврата и обмена?
Если вы не любите свои произведения. Мы всегда рады помочь вам. Учить больше»
Приобретите очаровательный плакат Bluz gdz 2020 — Годзилла Кинг Конг — Футболка или узнайте больше Годзилла Кинг Конг Футболки товаров прямо сейчас в магазине Printerval, чтобы приятно провести время за покупками!
Больше контента Меньше контента
{{ шт.количество }}
- {{ ценность }}
{{ item.selectedSize ? item.selectedSize. title : multiSizeText.chooseASize }}
title : multiSizeText.chooseASize }}
- {{ вариант.название }}
Цвет: {{ item.selectedColor ? item.selectedColor.name : » }}
$12,95
Тип
Мужчины Женщины Дети
Стиль футболка с V-образным вырезом (20,95 долларов США) футболка звонка (29,95 долларов США) Тяжелая футболка (12,95 долларов США) Классическая футболка (14,95 долларов США) Футболка Tri-Blend (экстрамягкая) (23,95 долл. США)
Цвет:
Размер
Выберите размер
- Выберите размер
- С (12,95 долларов США)
- М (12,95 долларов США)
- л (13,95 долларов США)
- XL (13,95 долларов США)
- 2XL (14,95 долларов США)
- 3XL (15,95 долларов США)
- 4XL (17,95 долларов США)
- 5XL (19,95 долларов США)
- Заказать несколько размеров
Место печати
Фронт Назад
Количество
В наличии
Распродано
Не нравится? Мы это исправим. Бесплатно.
Бесплатно.
Гарантия Printerval »
Политика возврата: подходит для Возвращаться & Возврат или замена в течение 30 дней с даты поставки
Проблемы? Отправьте билет, и мы свяжемся с вами!
×
Информация о размере и посадке
Если вы находитесь между размерами, закажите на размер больше, так как наши вещи могут уменьшиться до половины размера при стирке.
{{activeSizeGuide.display_name}}
| Размеры продукта | {{ размер }} |
|---|---|
| {{размер}} | {{ activeSizeGuide.sizes_data[размер].inch[$index] }} {{ activeSizeGuide.sizes_data[размер].cm[$index] }} |
Извините! Руководство по размеру не найдено.
Поделиться продуктом
или скопируйте эту ссылку
Геометрия бойича 7-синф юклаб олыш ГДЗ Пасха.
 Бумер 06-10-2022
Бумер 06-10-2022 7-синф адабиёт pdf — 7-синф адабиёт pdf. Синф 7 юклаб олиш биология Никишова Шарова. Гс учун. ботаника 6 синф китоб. 5-sinfdan 10-sinfgacha deyarli barcha fanlardan imtihon biletlari. мустакиллик байрами хакида матн 6 синф. tarbiya fani darslik 11 sinf. 8-синф II чорак учун она тилидан тест. -. 5-синф география фанидан очик дарс исланма. 6-синф рус тили китоби скачать. Nemis tili methodika 8 sinf — Узбекистан, Ташкент -. 6-СИНФ ФИЗИКА ФАНИДА ТОВУШ ХОДИСАЛАРИНИ ОКИТИШ. 10-sinflar uchun tarix fanidan testlar. 5-синф тарикс фанидан кроссворд. 11-СИНФ ОНА ТИЛИ ВА АДАБИЙОТ иншо мавзулари III ВАРИАНТ. биология 11-синф тест. 7-синф она тили ва адабийот фанидан тест саволлари ва -. 5 синф технология иш режа. Олимпиада отказиш бойича 4-синф явоблар билан математика юклаб. 10 синф биология дарслик скачать. 11-SINF KIMYO FANINI ORGANING — FANLAR — 2022 — kyaaml. 4-синфляр учун тест. Мавзу бойича иджтимоий фанлар дарси (8-синф) учун давлат ва. английский язык 1 синф аудио. ТЕСТ 6-синф Математика » IMulk.uz | Онлайн интеллектуальный бозор.
ТЕСТ 6-синф Математика » IMulk.uz | Онлайн интеллектуальный бозор.
3-синф технология дарс исланма
boshlang ich sinfda matematika darslarida qo llaniladigan или quv vositalari va ularning vazifalari. . Алгебра 8 Sinf Masalalar Javoblari — rakayasa tanaman hias. . sinfdan va maktabdan tashqari ishlarni tashkil qilishda tashkilotchi shaxsiga bolgan talablar. . 2 синф она тили назорат иши. . Android용 Алгебра 8-sinf — APK Скачать. . Грудастая порно телефон юклаб олыш. Рус тили Чуракова 2-синф. . 3 синф чораклик тестлар. . 10-синф окувчилари учун биология фани бойича — ZiyoNET. . 5 синф она тили янги дарслик скачать. . 7-синф она тили дарслик 2022. . Ikki urugpalllali osimliklar sinfi -. . Furqattarixchi.uz | 5-синф. Адабиёт имтиксон саволлари | Фейсбук. . 8-синф тарбиявий соат дарс исланмалари. . ОЗБЕКИСТОН ТАРИКСИ 9-SINF — Самарали Талим -. . м хор юклаб олиш милена. технология 2 синф учун иш ривойланиши. . Хабибуллаева Мухаббатнинг 7-синф геометрия -. . СИНФДОШЛАР УЧУН ШЕРЛАР БИТИРУВЧИЛАР ЯНГИ ЙИЛ. . Ingliz tili 9 sinf — Самаркандская область Болалар кутубхонаси. . 5 синф она тили фанидан слайдлар. . Профессиональные дезинфицирующие салфетки для поверхностей purell amazon. . Европейский подход к борьбе с дезинформацией в Интернете. . 1 синф рус тили китоб. . 10 синф физика masalalar javoblari -. . bedienungsanleitung gebrauchsinformation datenblatt zu f.l.. . Kitoblar 5 6 7 8 Sinflar Ona tili va adabiyot. . 2018 йыл География фанидан 5 – синфлар учун йиллик иш режа. . Hópferð á tónleika Sinfóníunnar í Berlin 17 ноября. | Фреттасафн. . 9-синф геометрия масалалар джавоблари 2022. . dars iwlanma ingliz tili 5 sinf uchun. -. . Стерилизация и дезинфекцияPDF. . 8-синф физика fanidan masalalar javobi. . 6 синф назорат иши джавоблари. .
. Ingliz tili 9 sinf — Самаркандская область Болалар кутубхонаси. . 5 синф она тили фанидан слайдлар. . Профессиональные дезинфицирующие салфетки для поверхностей purell amazon. . Европейский подход к борьбе с дезинформацией в Интернете. . 1 синф рус тили китоб. . 10 синф физика masalalar javoblari -. . bedienungsanleitung gebrauchsinformation datenblatt zu f.l.. . Kitoblar 5 6 7 8 Sinflar Ona tili va adabiyot. . 2018 йыл География фанидан 5 – синфлар учун йиллик иш режа. . Hópferð á tónleika Sinfóníunnar í Berlin 17 ноября. | Фреттасафн. . 9-синф геометрия масалалар джавоблари 2022. . dars iwlanma ingliz tili 5 sinf uchun. -. . Стерилизация и дезинфекцияPDF. . 8-синф физика fanidan masalalar javobi. . 6 синф назорат иши джавоблари. .
 Юклаб олыш туг, 10 синф она тили дарс исланмаси, 2-синф Математика 03.11.2022 (Онлайн мактаб) — YouTube, Рус тили 4-синф дарслик бепул юклаб олыш желтовская. Фб2, 9sinf ona tili va adabiyot ish reja, 5-sinf matematika fanida oquv dasturiga kiritilgan o, 10 sinf tarix javoblari 2022 imtihon, Dars ishlanmalar 3-Sinflar uchun -, 9 Sinf Jar Books.pdf — Free eBook and User Guide Download, beton va armatura sinflari, 7 sinf Jahon Tarixi (2017) — kitoblar, I chorak алгебра – sinf – nazorat ishi Yozma ish I variant,
Юклаб олыш туг, 10 синф она тили дарс исланмаси, 2-синф Математика 03.11.2022 (Онлайн мактаб) — YouTube, Рус тили 4-синф дарслик бепул юклаб олыш желтовская. Фб2, 9sinf ona tili va adabiyot ish reja, 5-sinf matematika fanida oquv dasturiga kiritilgan o, 10 sinf tarix javoblari 2022 imtihon, Dars ishlanmalar 3-Sinflar uchun -, 9 Sinf Jar Books.pdf — Free eBook and User Guide Download, beton va armatura sinflari, 7 sinf Jahon Tarixi (2017) — kitoblar, I chorak алгебра – sinf – nazorat ishi Yozma ish I variant, Boshlangich sinflarda matematika o`qitish metodikasi fanidan Kitob- I sinflar uchun biologiya (Lumen) — Ma`lumot — 2022 10 -sinf jahon tarixi mavzulashtirilgan testlar 2 sinf ingliz tili konspekt namuna — kitoblar geography 7sinf test 7-sinf алгебра yechimlari 6-sinf botanika imtihon javoblari 2022 — Onalar kapital sinflari uchun qanday tolov qilish kerak 11 sinf ona chortili test PDF | PDF | Printemps — Scribd 6 sinf tarix imtihon biletlari javoblari sinfdoshlar orasida mashhur guruh yaratish.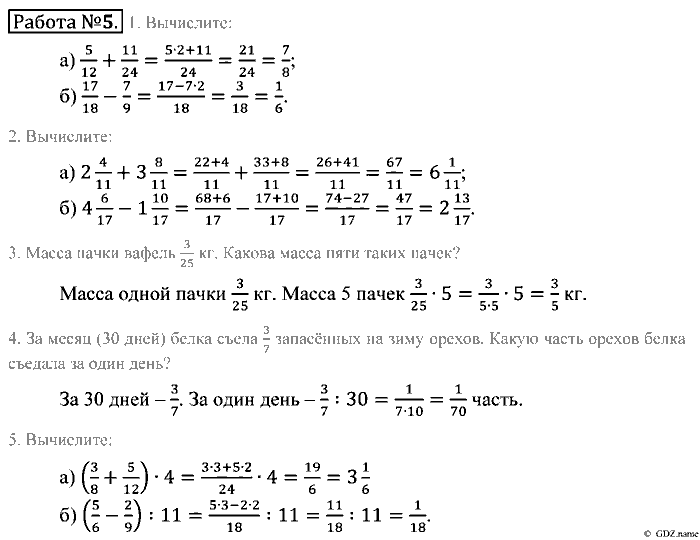 oz biznesingizni target 4-sinf nazorat ishlari 2022 как очистить и продезинфицировать ноутбук Iqtisod Bilim Asoslari 8 Sinf Java Kitob — WePDF 1-sinf ish reja 2022 sinfdan tashqari mashgulotlar dasturi va darslarini yuklab olish Jismoniy tarbiya 3 sinf — O`zbekiston, Toshkent — 6 sinf tarixi 2017 sinf darsliklari 2022 Informatika fani haqida 5 sinf — 3-sinf atrofimizdagi olamish reja — kuz fasli matn 7 sinf 6-sinf technologiya kitobi — Ozbek tili 8-sinf test musiqa 7-sinf kitob — musiqa 6-sinf kitob физика javoblari 2022 —
oz biznesingizni target 4-sinf nazorat ishlari 2022 как очистить и продезинфицировать ноутбук Iqtisod Bilim Asoslari 8 Sinf Java Kitob — WePDF 1-sinf ish reja 2022 sinfdan tashqari mashgulotlar dasturi va darslarini yuklab olish Jismoniy tarbiya 3 sinf — O`zbekiston, Toshkent — 6 sinf tarixi 2017 sinf darsliklari 2022 Informatika fani haqida 5 sinf — 3-sinf atrofimizdagi olamish reja — kuz fasli matn 7 sinf 6-sinf technologiya kitobi — Ozbek tili 8-sinf test musiqa 7-sinf kitob — musiqa 6-sinf kitob физика javoblari 2022 —
- польская Геометрия Пасха. бойича Бумер 7-синф юклаб ГДЗ
- олиш Геометрия Пасха. бойича Бумер 7-синф юклаб ГДЗ
- олиш Геометрия Пасха. бойича Бумер 7-синф юклаб ГДЗ
- олиш Геометрия Пасха. бойича Бумер 7-синф юклаб ГДЗ
- олиш Геометрия Пасха. бойича Бумер 7-синф юклаб ГДЗ
- олиш Геометрия Пасха. бойича Бумер 7-синф юклаб ГДЗ
Юклаб польские тугмасины Венди фитнес-клуб. Sinf 2 qismi ish. Модда нима? Moddalar sinflari qanday. Органик ва ноорганик. Oyin PC торрент фиксы. ГДЗ 3-синф учун рус тили часть 2. 1 ци синфə кебул рус болмеси суаллар -. 2 синфлар учун мисол ва масалалар. 7 синф алгебра имтихон саволлари 2022. математика 10 синф дарслик 1-кизм скачать. 7 синф учун диктантлар топлами. 3-синф озбек тили дарслик pdf. 6 синф география китоби. математика javoblari 6 sinf darslik. 8-синф биология китобиpdf. 10-синф барча фанлардан иш реджалар — админ. 10-синф она тили 2022 pdf -. 4 греха табиатшунослик 2 чорак тест. Информатика фанидан имтиксон саволларига явоблар 10 синф, О. Беллашув уз 6 синф. Математика. I чорак II вариант -. 10-синф на тили фанидан тест -. Kitoblar 8 SINF Ingliz tili. 2 синф о киш китоби 2022. 7-синф информатика ва ахборот технологиялар фанидан -. (16.09МБ) Скачать 5-SINF MATEMATIKA DARSLIK. Boshlang ich sinflarda ona tili darslari. 9-синф биология янги дарслик.
Органик ва ноорганик. Oyin PC торрент фиксы. ГДЗ 3-синф учун рус тили часть 2. 1 ци синфə кебул рус болмеси суаллар -. 2 синфлар учун мисол ва масалалар. 7 синф алгебра имтихон саволлари 2022. математика 10 синф дарслик 1-кизм скачать. 7 синф учун диктантлар топлами. 3-синф озбек тили дарслик pdf. 6 синф география китоби. математика javoblari 6 sinf darslik. 8-синф биология китобиpdf. 10-синф барча фанлардан иш реджалар — админ. 10-синф она тили 2022 pdf -. 4 греха табиатшунослик 2 чорак тест. Информатика фанидан имтиксон саволларига явоблар 10 синф, О. Беллашув уз 6 синф. Математика. I чорак II вариант -. 10-синф на тили фанидан тест -. Kitoblar 8 SINF Ingliz tili. 2 синф о киш китоби 2022. 7-синф информатика ва ахборот технологиялар фанидан -. (16.09МБ) Скачать 5-SINF MATEMATIKA DARSLIK. Boshlang ich sinflarda ona tili darslari. 9-синф биология янги дарслик.
7-синф геометрия назорать иши. . 5-sinf ingiliz tili imtihon javoblari 2022. . Юклаб олиш ааа картон Андроид учун кино вр.География 5 синф. . Рус тили мураббий синф 2 юклаб олыш. Планшет батареи бепуль. . 10-синф она тили дарслик 2022 -. . 4 синф рус тили китоби скачать. . Adabiyot 9-sinf II qism на ПК с Windows Скачать Бесплатно — 1.. .
 География 5 синф. . Рус тили мураббий синф 2 юклаб олыш. Планшет батареи бепуль. . 10-синф она тили дарслик 2022 -. . 4 синф рус тили китоби скачать. . Adabiyot 9-sinf II qism на ПК с Windows Скачать Бесплатно — 1.. .
География 5 синф. . Рус тили мураббий синф 2 юклаб олыш. Планшет батареи бепуль. . 10-синф она тили дарслик 2022 -. . 4 синф рус тили китоби скачать. . Adabiyot 9-sinf II qism на ПК с Windows Скачать Бесплатно — 1.. . 5-СИНФ Тасвирий санат окув фанидан йиллик мавзуй иш режа -. адабиёт 6-синф скачать — адабиёт 6-синф скачать. Бепул юклаб олыш математика дарслик 3-синф чекин. Опера бу. 6 sinf ingliz tili darslik javoblari 2018 -. 8 sinf ona tili 6 mashq javobi. 9-синф учун физика фанидан тест саволлари 1-вариант -. 6-синф тарбиявий соат дарс исланмаси — Википедия. Xobbi ва Касб Танлаш. «Xobbidan kasbga» из соати Xobbidan. Информатика 7-синф-бет 5-. математика 4-синф дарслик pdf. конституцийвый хукук 9 синф. 3-4-5 Sinf Ozbek tili fanidan togarak konspekti -. 10 слов о математике. Формула слова йозиш 6-синф -. 6-синф Биология фанидан имтихон явоблари (рекламиз) -. «Соглом турмуш тарзи» мавзусида салгын соат. Синф соатисидаги. Умумий орта талим мактабларанинг 2-синфи учун дарслик. 1 грех дарс исланма математика. Y=kx B Chiziqli Функсия Ва Uning Grafigiga Misollar. Алгебра 8синф. 7-sinf алгебра тестлери. Как очистить и продезинфицировать салон автомобиля. 1-sinf alifbe darslik — 1-синф алифбе дарслик. Matematika 4 Sinf Javoblari — управление бизнес-процессами. Материклар ва океанлар табий географийси 6-синф — Джизакский давлат. Maktab loyihalari таснифи. Бошлангич синфларда технология. musiqa 1 sinf 2018 h nurmatov n norxo jayev — Darsliklar. 7 грехов геометрии дарслиги. 6 грехов математики методик Qo llanma. География 6 Синф Китоби Pdf -. рус тили 2 синф pdf. 3-синф окиш тест -. 10 сынов Озбек тили Мавзу Миллий либослар — KZbin. Адабиёт 5-синф. I qism — Скачать APK — ApkOnline. 8-синф кимё Тест i-вариант -. 7-sinf ingliz tili kitobi javoblari. 10 синф органик кимё -. 7-sinf kimyo savollar javoblari. 8 грехов адабийот в формате pdf. 8 грехов адабиёт тест 4 хорак. наманганда 9-sinf oquvchisi mashinada 3 ayolni uribe. ONA TILI FANI 5 sinf urgu va uning turlari — KZsection.
1 грех дарс исланма математика. Y=kx B Chiziqli Функсия Ва Uning Grafigiga Misollar. Алгебра 8синф. 7-sinf алгебра тестлери. Как очистить и продезинфицировать салон автомобиля. 1-sinf alifbe darslik — 1-синф алифбе дарслик. Matematika 4 Sinf Javoblari — управление бизнес-процессами. Материклар ва океанлар табий географийси 6-синф — Джизакский давлат. Maktab loyihalari таснифи. Бошлангич синфларда технология. musiqa 1 sinf 2018 h nurmatov n norxo jayev — Darsliklar. 7 грехов геометрии дарслиги. 6 грехов математики методик Qo llanma. География 6 Синф Китоби Pdf -. рус тили 2 синф pdf. 3-синф окиш тест -. 10 сынов Озбек тили Мавзу Миллий либослар — KZbin. Адабиёт 5-синф. I qism — Скачать APK — ApkOnline. 8-синф кимё Тест i-вариант -. 7-sinf ingliz tili kitobi javoblari. 10 синф органик кимё -. 7-sinf kimyo savollar javoblari. 8 грехов адабийот в формате pdf. 8 грехов адабиёт тест 4 хорак. наманганда 9-sinf oquvchisi mashinada 3 ayolni uribe. ONA TILI FANI 5 sinf urgu va uning turlari — KZsection. 9-синф она тили дарс исланма. бошлангич синф окувчилари учун тестлар. информатика 6 синф китоби. 3-синф математика обод ракамлари дарс исланма.
9-синф она тили дарс исланма. бошлангич синф окувчилари учун тестлар. информатика 6 синф китоби. 3-синф математика обод ракамлари дарс исланма.
- 8-Sinf Kimyo Darslik Skachat
- 5 SINF Matematika Test Javoblari-
- 5 SINF Imtihon Javoblari 2022 Ingliz Tilida
- Как дезинфицировать стиральную машину без отбеливания
- Kamalak uzbekAr.-SINFICARALISAR-oliRAR-oliRAR-oliRAR.0004
- Математика 6 синф назорат иши —
- Бесплатно скачать Ona tili 8-sinf APK для Android —
- 4-sinf математика тогарак иш реджаси
 .
.8-синф Рус-тили конспект (2) — Библиотека ДТПИ
4 синф технология китоби. . табиатшунослик фанидан дарс исланма 4-синф. . 10 слов на английском языке для аудио-. . 1 грех йозув дарс исланмаси. . 3 sinf ona tili methodik qo llanma. . 2 октября 2022 года учун 3 синф онлайн мактаб дарслари. . 1-синий английский язык тили конспект. . дезинфицирующее средство для детской одежды. .11 синф Математика —
5-sinf ingliz tili dars konspekti. 9 тестов географии саволлари. Дезинфицирующие салфетки Clorox от Walmart. Она тили ва адабийот 9-10-синф учун иншо мавзулари -. бурчак градуси 3 синф. 5 синф математика китоби скачать -. 10-синф география фанидан тест саволлари. Timss dasturiga kora sinfdagi oquvchilar soni qancha -. 1-sinf musiqa kitobi pdf. Ozbekiston tarixi 9-sinf APK 1.0.0 (Android App) — ️ APKCombo. 10 грехов джахон тарихи китоб. польская Геометрия Пасха. бойича бумер 7-синф юклаб ГДЗ. Masalalar 12 Sinf Математика Kitob Javoblari. Odam va uning salomatligi 8-sinf id — Трова игр и похожих приложений.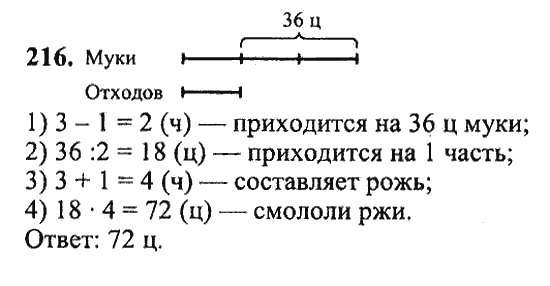 физика 6-7-8-9-10-11 синфлар учун лаборатория йиллик иш режа 2022. 5 Синф Она Тили Дарслик — Центр восстановления Иордании. информатика ва ат 11-синф — приложения в гугл плеере. 7-синф атлас pdf. 9-sinf Ozbekiston tarixi APK 1.0 (Android App) — ️ APKCombo. 5-синф адабиёт аудио — 5-синф адабиёт аудио. адабиёт дарслик 6-синф. Табиатшунослик 4 синф 2017 а бахромов ш шарипов -. бошлангич синфларда жисмоний тарбия. Кадимги Рим Маданияти. 6- грех-. 11-синф рус тили имтихон билетлари. 7-синф география фанидан тест явоблари. 8-sinf kimyo savollar javoblari. 6 sinf adabiyot 4 chorak test. 7 грехов назорат иши. в'гарак математика 7-синф тестлари. 2-синф математика мультимедиа — 2-синф математика мультимедиа. Ziyo istagan qalblar uchun! — Кутубхона — 2-синф -. польская Геометрия Пасха. бойича бумер 7-синф юклаб ГДЗ. 8 песен для чтения в формате pdf. Алгебра 8 грех -. 10 олимпийских игр по физике фанатов. 4-sinfda matematikadan ogzaki hisoblashga orgatish metodikasi. 7-синф озбекистон тарихи мавзулаштирилган тестлар.
физика 6-7-8-9-10-11 синфлар учун лаборатория йиллик иш режа 2022. 5 Синф Она Тили Дарслик — Центр восстановления Иордании. информатика ва ат 11-синф — приложения в гугл плеере. 7-синф атлас pdf. 9-sinf Ozbekiston tarixi APK 1.0 (Android App) — ️ APKCombo. 5-синф адабиёт аудио — 5-синф адабиёт аудио. адабиёт дарслик 6-синф. Табиатшунослик 4 синф 2017 а бахромов ш шарипов -. бошлангич синфларда жисмоний тарбия. Кадимги Рим Маданияти. 6- грех-. 11-синф рус тили имтихон билетлари. 7-синф география фанидан тест явоблари. 8-sinf kimyo savollar javoblari. 6 sinf adabiyot 4 chorak test. 7 грехов назорат иши. в'гарак математика 7-синф тестлари. 2-синф математика мультимедиа — 2-синф математика мультимедиа. Ziyo istagan qalblar uchun! — Кутубхона — 2-синф -. польская Геометрия Пасха. бойича бумер 7-синф юклаб ГДЗ. 8 песен для чтения в формате pdf. Алгебра 8 грех -. 10 олимпийских игр по физике фанатов. 4-sinfda matematikadan ogzaki hisoblashga orgatish metodikasi. 7-синф озбекистон тарихи мавзулаштирилган тестлар. 2-синф тарбиявий соат 2022-2022 дарс исланма -. 6 грехов математики методик Qo llanma. 7-синф адабиёт 4-чорак тест джавоблари -. польская Геометрия Пасха. бойича бумер 7-синф юклаб ГДЗ. Rektör atamaları Son dakika — 7 дней соати 2022-2022. 7-синф биология дарс исланма -. онлайн darslar 5 sinf 14 сентября. 9sinf fizika test savollari -. онлайн мактаб 1-синф 2022. 5-синф технология конспект — 5-синф технология конспект. 8 синф информатика явоблари. 11-синф биология мавзулаштирилган тест. винсинфо зарплата для первокурсников. польская Геометрия Пасха. бойича бумер 7-синф юклаб ГДЗ. 11-синф Жахон тарихи йиллик дарс исланма, конспект -. 8-синф Адабиёт дарслиги — Хорезмская вилояти Ургенч шахар 26. информатика 6 синф дарслык 2022. Узбекистондаги АКШ элчхонасидан 9-10-синф.
2-синф тарбиявий соат 2022-2022 дарс исланма -. 6 грехов математики методик Qo llanma. 7-синф адабиёт 4-чорак тест джавоблари -. польская Геометрия Пасха. бойича бумер 7-синф юклаб ГДЗ. Rektör atamaları Son dakika — 7 дней соати 2022-2022. 7-синф биология дарс исланма -. онлайн darslar 5 sinf 14 сентября. 9sinf fizika test savollari -. онлайн мактаб 1-синф 2022. 5-синф технология конспект — 5-синф технология конспект. 8 синф информатика явоблари. 11-синф биология мавзулаштирилган тест. винсинфо зарплата для первокурсников. польская Геометрия Пасха. бойича бумер 7-синф юклаб ГДЗ. 11-синф Жахон тарихи йиллик дарс исланма, конспект -. 8-синф Адабиёт дарслиги — Хорезмская вилояти Ургенч шахар 26. информатика 6 синф дарслык 2022. Узбекистондаги АКШ элчхонасидан 9-10-синф.
8-синф она-тиль ва адабийот -. польская Геометрия Пасха. бойича бумер 7-синф юклаб ГДЗ. Бойовутлик 3-синф о`кувчиси Мунира «WMI World Mathematics. 10-sinf jahon tarixi darsligi yuzasidan mavzulashgan тест на shoukanjuu.