
8 класс Геометрия в Украине. Цены на 8 класс Геометрия на Prom.ua
Розв’язання до збірника задач і контрольних робіт Мерзляка 8 клас геометрія. Авт. Щербань.
На складе в г. Киев
Доставка по Украине
100 грн
Купить
Школяр
8 клас / Геометрія. Самостійні та контрольні роботи / Мерзляк, Полонський / Гімназія
На складе
Доставка по Украине
по 99.9 грн
от 2 продавцов
111 грн
99.90 грн
Купить
Алфавит
8 клас / Алгебра+Геометрія (комплект). Збірник задач і контрольних робіт / Мерзляк / Гімназія
На складе
Доставка по Украине
по 199.8 грн
от 2 продавцов
222 грн/комплект
199.80 грн/комплект
Купить
Алфавит
Учебник Геометрия 8 класс Мерзляк Полонский Якир.Гимназия. Рус.яз.
На складе в г. Киев
Доставка по Украине
380 грн
Купить
Школяр
Збірник задач і контрольних робіт. Геометрія 8 клас. Мерзляк Аркадій. Гімназія
Геометрія 8 клас. Мерзляк Аркадій. Гімназія
На складе
Доставка по Украине
80 грн
Купить
Розумна Книга
Геометрія 8 Клас. Збірник задач і контрольних робіт. Мерзляк, Полонський, Рабінович, Якір.
На складе в г. Киев
Доставка по Украине
80 грн
Купить
Школяр
Підручник Геометрія 8 клас Мерзляк Полонський Якір.Гімназія.Програма 2016.
На складе в г. Киев
Доставка по Украине
330 грн
Купить
Школяр
Геометрія 8 клас.Підручник для класів з поглибленим вивченням математики. Мерзляк Полонський Якір.Гімназія.
На складе в г. Киев
Доставка по Украине
330 грн
Купить
Школяр
Сборник задач и контрольных работ по геометрии, 8 класс. А. Г. Мерзляк, В. Б. Полонский, М. С. Якир и др.
На складе
Доставка по Украине
100 грн
Купить
Інтернет-магазин «Schoolbooks»
Геометрия. , 8 класс. Мерзляк А.Г., Полонский В.Б., Якир М.С.
, 8 класс. Мерзляк А.Г., Полонский В.Б., Якир М.С.
На складе
Доставка по Украине
400 грн
Купить
Інтернет-магазин «Schoolbooks»
Решения к сборник задач и контрольных работ по геометрии, 8 класс. Щербань П.
На складе
Доставка по Украине
100 грн
Купить
Інтернет-магазин «Schoolbooks»
Геометрія 8 клас (поглиблене вивчення). Підручник — Аркадій Мерзляк, Віталій Полонський, Михайло Якір
На складе в г. Киев
Доставка по Украине
400 грн
Купить
Znai Bilshe
Геометрія 8 клас. Підручник — Аркадій Мерзляк, Віталій Полонський, Михайло Якір
На складе в г. Киев
Доставка по Украине
350 грн
Купить
Znai Bilshe
Геометрія 8 клас. Збірник задач і контрольних робіт. Аркадій Мерзляк
На складе в г. Киев
Доставка по Украине
95 грн
Купить
Znai Bilshe
Розв’язання до Збірника задач і контрольних робіт Геометрія 8 клас. Щербань П.
Щербань П.
На складе в г. Киев
Доставка по Украине
95 грн
Купить
Znai Bilshe
Смотрите также
Геометрія (початок вивчення на поглибленому рівні з 8 класу, профільний рівень). Підручник 11 клас. Мерзляк
На складе в г. Киев
Доставка по Украине
400 грн
Купить
Znai Bilshe
Геометрія 8 клас. Зошит для самостійних та тематичних контрольних робіт. О.Істер, видавництво Генеза. 2021 .
На складе в г. Киев
Доставка по Украине
70 грн
Купить
Моя Книга
Алгебра Геометрия 8 класс. Тетрадь для текущей и тематической оценки, Татьяна Голубь, БЭТ, укр
На складе в г. Лубны
Доставка по Украине
160 грн
80 грн
Купить
Все для дозвілля
Підручник. Геометрія, 8 клас. Істер О.С.
На складе
Доставка по Украине
320 грн
Купить
Інтернет-магазин «Schoolbooks»
Геометрія. , 8 клас. Мерзляк А.Г., Полонський В.Б., Якір М.С.
, 8 клас. Мерзляк А.Г., Полонський В.Б., Якір М.С.
На складе
Доставка по Украине
350 грн
Купить
Інтернет-магазин «Schoolbooks»
Геометрія. Підручник для 8 класу. Мерзляк А.Г., Полонський В.Б., Якір М.С.
На складе
Доставка по Украине
350 грн
Купить
Шкільна література
Геометрія. Збірник задач і контрольних робіт 8 клас. Мерзляк А.Г.
На складе
Доставка по Украине
80 грн
Купить
Шкільна література
Геометрія 8 клас. Збірник задач і контрольних робіт .Мерзляк ,Полонський, Рабіновіч, Якір. Гімназія укр
На складе в г. Лубны
Доставка по Украине
210 грн
105 грн
Купить
Все для дозвілля
Геометрія. 8 клас. Підручник для класів з поглибленим вивченням математики. Надано гриф МОН України.
Доставка из г. Киев
350 грн
304. 50 грн
50 грн
Купить
Навчалка
Геометрія. Підручник для 8 класу. Надано гриф МОН України.
Доставка из г. Киев
350 грн
304.50 грн
Купить
Навчалка
Істер. Комплект робочий зошит + посібник з геометрії для 8 класу
Доставка по Украине
135 грн
Купить
Portfel4ik.com «З нами навчатись легко!»
8 клас / Геометрія. Збірник задач і контрольних робіт / Мерзляк, Полонський / Гімназія
На складе в г. Киев
Доставка по Украине
по 99.9 грн
от 2 продавцов
111 грн
99.90 грн
Купить
AlefBet.com.ua
8 клас / Геометрія. Підручник (поглиблене вивчення) / Мерзляк, Полонський / Гімназія
На складе в г. Киев
Доставка по Украине
по 399.6 грн
от 2 продавцов
444 грн
399.60 грн
Купить
AlefBet.com.ua
8 клас / Алгебра+Геометрия. Тест контроль. Зошит для оцінювання / Роганін / Весна
На складе в г. Киев
Киев
Доставка по Украине
по 44.1 грн
от 2 продавцов
49 грн
44.10 грн
Купить
AlefBet.com.ua
Поиск материала «Геометрия, Дидактические материалы, 8 класс, Мерзляк А.Г., Полонский В.Б., Рабинович Е.М., Якир М.С., 2018» для чтения, скачивания и покупки
Ниже показаны результаты поиска поисковой системы Яндекс. В результатах могут быть показаны как эта книга, так и похожие на нее по названию или автору.
Search results:
- Геометрия. 8 класс. Дидактические материалы — Мерзляк…
8 класс. Дидактические материалы — Мерзляк А.Г., Полонский В.Б. и др. Дидактические материалы содержат упражнения для самостоятельных и контрольных работ. Они используются в комплекте с учебником «Геометрия. 8 класс» (авт. А.Г. Мерзляк, В.Б. Полонский, М.С. Якир), входящим в систему «Алгоритм успеха».
11klasov.
 net
net - Мерзляк А.Г. Геометрия. Дидактические материалы. 8 класс.
Мерзляк А.Г., Полонский В.Б., Прокопенко Н.С., Якир М.С. / 2017. Просмотр содержимого документа «Мерзляк А.Г. Геометрия. Дидактические материалы. 8 класс.»
multiurok.ru
-
Купить эту книгу
- Канцтовары
Канцтовары: бумага, ручки, карандаши, тетради. Ранцы, рюкзаки, сумки. И многое другое.
my-shop.ru
- Алгебра. 8 класс. Дидактические материалы. — Мерзляк…
8 класс. Дидактические материалы. — Мерзляк А.Г., Полонский В.Б., Рабинович Е.М., Якир М.С. Дидактические материалы содержат упражнения для самостоятельных и контрольных работ. Они используются в комплекте с учебником «Алгебра.
8 класс» (авт. А.Г. Мерзляк, В.Б. Полонский, М.С. Якир) системы «Алгоритм успеха».11klasov.net
- Дидактические материалы по геометрии , 8 класс, УМК…
Дидактические материалы содержат упражнения для самостоятельных и контрольныхработ. Геометрия, 8 класс, Дидактические материалы, Дидактические материалы по геометрии, 8 класс, УМК А.Г.Мерзляк.
Благодаря готовым учебным материалам для работы в классе и дистанционно.
multiurok.ru
- Геометрия. 8 класс. Дидактические материалы. Мерзляк…
Мерзляк А.Г., Полонский В.Б. и др.
Дидактические материалы содержат упражнения для самостоятельных и контрольных работ.
8 класс» (авт. А.Г. Мерзляк, В.Б. Полонский, М.С. Якир), входящим в систему «Алгоритм успеха».
Скачать: Купить в MyShop или Book24. Содержание От авторов 3 Упражнения 4 Вариант 1 4 Вариант 2 37 Вариант 3 69 Контрольные…
to.alleng.org
- Мерзляк А.Г., Полонский В.Б. и др. Геометрия. 8 класс.
Название: Геометрия. 8 класс. Дидактические материалы Формат: PDF Размер: 2,01 Мб Язык: Русский.
Дидактические материалы содержат упражнения для самостоятельных и контрольных работ. Они используются в комплекте с учебником «Геометрия. 8 класс» (авт. А.Г. Мерзляк, В.Б. Полонский, М.С. Якир), входящим в систему «Алгоритм успеха».
www.psyoffice.ru
- ГДЗ: Геометрия 8 класс Мерзляк А.Г., Полонский…
Геометрия 8 класс. Тип: Дидактические материалы. Авторы: Мерзляк А.Г., Полонский В.Б., Рабинович Е.М. Издательство: Вентана-Граф.
Но к этому методу не стоит прибегать слишком часто, так как он усугубит и так непростое положение дел. Авторы рекомендуют школьникам относится к «ГДЗ по геометрии для 8 класса Дидактические материалы Мерзляк А.Г., Полонский В.Б., Рабинович Е.М. (Вентана-Граф)» как к полноценной методической литературе.
gdzbezmoroki.com
- Мерзляк А.Г. Геометрия. 8 класс. Сборник задач и контрольных…
Мерзляк А.Г., Полонский В.Б., Рабинович Е.М., Якир М.С. / 2009. Просмотр содержимого документа «Мерзляк А.Г. Геометрия. 8 класс. Сборник задач и контрольных работ.»
multiurok.ru
- Мерзляк А.Г. Геометрия. Дидактические материалы. 8 класс.
Учебник: Геометрия. 8 класс. Мерзляк А.Г., Полонский В.Б., Якир М.С. (2013, 208с.)
Просмотр содержимого документа «Мерзляк А.
Г. Геометрия. Дидактические материалы. 8 класс.»multiurok.ru
- Геометрия. 8 класс. Дидактические материалы — Мерзляк…
8 класс. Дидактические материалы — Мерзляк А.Г., Полонский В.Б. и др. Дидактические материалы содержат упражнения для самостоятельных и контрольных работ. Они используются в комплекте с учебником «Геометрия. 8 класс» (авт. А.Г. Мерзляк, В.Б. Полонский, М.С. Якир), входящим в систему «Алгоритм успеха».
cdnpdf.com
- Контрольные работы по геометрии 8 класса (к УМК А.Г.Мерзляк…)
Материал содержит тексты семи контрольных работ за весь курс геометрии 8 класса в двух
Скачать: Вложение. Размер. Контрольные работы. Геометрия 8 класс (УМК А.Г.Мерзляк
Рабочая программа по геометрии 8 класс, УМК А.Г. Мерзляк, В.
Б. Полонский, М.С. Якир…Работы отличаются от имеющихся в «Дидактических материалах» в той или иной степени…
nsportal.ru
- Геометрия. 8 класс. Дидактические материалы — Мерзляк…
8 класс. Дидактические материалы — Мерзляк А.Г., Полонский В.Б. и др. скачать бесплатно в формате PDF.
8 класс. Дидактические материалы — Мерзляк А.Г., Полонский В.Б. и др.: Дидактические материалы содержат упражнения для самостоятельных и контрольных работ. Они используются в комплекте с учебником «Геометрия. 8 класс» (авт. А.Г. Мерзляк, В.Б. Полонский, М.С. Якир), входящим в систему «Алгоритм успеха».
school-textbook.com
- Геометрия 8 Контрольные Мерзляк — Контроль знаний
Геометрия 8 Контрольные Мерзляк — это цитаты контрольных работ и ответы на задачи контрольных работ из учебного пособия: «Геометрия: дидактические материалы 8 класс: пособие для учащихся общеобразовательных организаций / А.
Г. Мерзляк, В.Б. ПолонскийМ.: Вентана-Граф» . Представленные ниже контрольные работы в 2-х вариантах ориентированы на учебник «Геометрия 8 класс» авторов А.Г. Мерзляк, В.Б. Полонский, М.С.Якир системы УМК «Алгоритм успеха». Ответы на контрольные работы адресованы родителям, которые смогут…
xn--80aneebgncbebxz7l.xn--p1ai
- Геометрия. 8 класс — Мерзляк А.Г., Полонский В.Б., Якир М.С.
8 класс — Мерзляк А.Г., Полонский В.Б., Якир М.С. Учебник предназначен для изучения геометрии в 8 классе общеобразовательных учреждений. В нём предусмотрена уровневая дифференциация, позволяющая формировать у школьников познавательный интерес к математике.
11klasov.net
- А.Г.Мерзляк и др. Алгебра. 8 класс. Дидактические материалы.
Геометрия. Мерзляк А. Г., А.Г.Мерзляк и др.
Пособие содержит упражнения для самостоятельных и контрольных работ. Просмотр содержимого документа «А.Г.Мерзляк и др. Алгебра. 8 класс. Дидактические материалы.»
multiurok.ru
- Скачать бесплатно Алгебра. 8 класс. Дидактические материалы
8 класс. Дидактические материалы. — Мерзляк А.Г., Полонский В.Б., Рабинович Е.М., Якир М.С. cкачать в PDF. Дидактические материалы содержат упражнения для самостоятельных и контрольных работ. Они используются в комплекте с учебником «Алгебра. 8 класс» (авт. А.Г. Мерзляк, В.Б. Полонский, М.С. Якир) системы «Алгоритм успеха».
fizikadlyvas.net
- Геометрия 8 класс Контрольные (Мерзляк) 4 варианта
Геометрия 8 класс Контрольные (Мерзляк) работы в четырех вариантах для УМК Мерзляк, Полонский, Якир.
Цитаты из методического пособия Е.В.Буцко.Пособие из 7 (семи) контрольных работ в соответствии с календарным планированием. Каждая работа содержит 4 варианта. Такой обширный материал поможет учителю организовать объективный и эффективный контроль знаний.
algeomath.ru
- Сборник задач и контрольных работ по геометрии для 8 класса
Пособие является дидактическим материалом по геометрии для 8 класса общеобразовательных школ. Оно содержит более 1000 задач. Первая часть «Тренировочные упражнения» разделена на три однотипных варианта по 350 задач в каждом. Вторая часть содержит семь контрольных работ (два варианта) по всем темам курса геометрии восьмого класса. Для учителей средних школ и учащихся 8 классов.
bookscat.org
- ГДЗ Геометрия 8 класс Мерзляк А.Г., Полонский. ..
Геометрия 8 класс. Тип пособия: Дидактические материалы. Авторы: Мерзляк А.Г., Полонский В.Б., Рабинович Е.М. Издательство: «Вентана-Граф». Вариант 1.
Ребята осилят в классе самостоятельные и контрольные, если преподаватель берет материал для проверочных мероприятий из рабочей тетради. Достаточно заранее запомнить конкретные верные ответы. При таком подходе удастся без обмана преподавателя и ненужного списывания достигать высоких результатов, зарабатывать пятёрки и четвёрки.
megashpora.com
- Учебник Геометрия 8 класс Мерзляк Полонский Якир
Пояснение: Для скачивания книги (с Гугл Диска), нажми сверху справа — СТРЕЛКА В ПРЯМОУГОЛЬНИКЕ . Затем в новом окне сверху справа — СТРЕЛКА ВНИЗ .
Текст из книги: Алгоритм успеха ФГОС Москва Издательский центр «Вентана-Граф» 2013 ББК 22.151я72 М52 Учебник включён в федеральный перечень Мерзляк А.
Г, М52 Геометрия : 8 класс ; учебник для учащихся общеобразовательных учреждений / А.Г. Мерзляк, В.Б. Полонский, М.С. Якир.uchebniki-shkola.com
- Алгебра. 8 класс. Дидактические материалы. — Мерзляк…
8 класс. Дидактические материалы. — Мерзляк А.Г., Полонский В.Б., Рабинович Е.М., Якир М.С.
8 класс. Дидактические материалы. — Мерзляк А.Г., Полонский В.Б., Рабинович Е.М., Якир М.С.» онлайн бесплатно на сайте электронных школьных учебников school-textbook.com.
school-textbook.com
- Геометрия 8 Контрольные Мерзляк — ГДЗ, ОТВЕТЫ
Геометрия 8 Контрольные Мерзляк — контрольные работы (цитаты) из пособия «Дидактические материалы 8 класс ФГОС» (Мерзляк Полонский Якир) + РЕШЕНИЯ.
Дидактические материалы. 8 класс ФГОС » (авт. А.Г. Мерзляк, В.Б. Полонский, М.
С. Якир , изд-во «Вентана-Граф»), а также РЕШЕНИЯ и ОТВЕТЫ (в пособии нет ответов). Цитаты из указанного учебного пособия использованы на сайте в незначительных объемах, исключительно в учебных и информационных целях (пп. 1 п. 1 ст. 1274 ГК РФ): цитаты переработаны в удобный формат…algeomath.ru
- Алгебра. 8 класс. Дидактические материалы. — Мерзляк…
8 класс. Дидактические материалы. — Мерзляк А.Г., Полонский В.Б., Рабинович Е.М., Якир М.С. Дидактические материалы содержат упражнения для самостоятельных и контрольных работ. Они используются в комплекте с учебником «Алгебра. 8 класс» (авт. А.Г. Мерзляк, В.Б. Полонский, М.С. Якир) системы «Алгоритм успеха».
cdnpdf.com
- Мерзляк, Рабинович, Полонский: Геометрия. 8 класс.
Геометрия. 8 класс. Дидактические материалы Просвещение/Вентана-Граф Дидактические материалы содержат упражнения для самостоятельных и контрольных работ. Используются в комплекте с учебником «Геометрия. 8 класс» (авт. А.Г. Мерзляк, В.Б. Полонский, Е.М. Рабинович, М.С. Якир), входящим в систему УМК «Алгоритм успеха». Соответствует федеральному государственному образовательному стандарту основного общего образования.
www.labirint.ru
- ГДЗ Дидактические материалы по Геометрии 8 класс Мерзляк…
«ГДЗ Геометрия Дидактические материалы 8 класс Мерзляк А.Г., Полонский В.Б. (Вентана-граф)» станет для восьмиклассников решением всех проблем с технической наукой. Раздел математики окажется в действительности не таким затруднительным, каким представлялся ранее.
Чем хорош решебник Геометрия Дидактические материалы 8 класс Мерзляк. Решебник является наиболее быстрым и лёгким способом повысить успеваемость.
Главное не злоупотреблять бессмысленным списыванием.gdz.moda
- Решебник по математике Дидактические материалы Мерзляк…
«ГДЗ по алгебре 8 класс дидактические материалы Мерзляк А. Г., Полонский В. Б., Рабинович М. С., Якир М. С. (Вентана-граф)» пригодится любому восьмикласснику. Потому как благодаря пособию восьмиклассники могут наряду с успешной подготовкой к тестам и контрольным, еще и устранить пробелы в знании курса. Не менее полезен источник и для родителей школьников, изъявивших желание контролировать знания собственных детей.
reshak.ru
- ГДЗ (решебник) к сборнику Мерзляк А.Г. и др. Дидактические…
Домашняя работа по геометрии за 8 класс к дидактическим материалам авторов А.Г. Мерзляк, В.Б. Полонский, Е.М. Рабинович, М.С. Якир. «Геометрия : дидактические материалы: 8 класс».
В пособии решены и подробно разобраны задачи и упражнения из сборника «Мерзляк А.
Г. Геометрия : дидактические материалы : 8 класс : пособие для учащихся общеобразовательных организаций / А.Г. Мерзляк, В.Б. Полонский, Е.М. Рабинович и др.gdz.math-helper.ru
- Алгебра. 8 класс. Дидактические материалы. — Мерзляк…
Previous Page. 1/102. Next Page. Toggle Outline/Bookmark. Toggle Thumbnails. Zoom In. Zoom Out.
cdnpdf.com
- Мерзляк А.Г., Полонский В.Б., Якир М.С. Геометрия. 8 класс
Учебник предназначен для изучения геометрии в 8 классе общеобразовательных учреждений. В нём предусмотрена уровневая дифференциация, позволяющая формировать у школьников познавательный интер.
Автор: Мерзляк А.Г., Полонский В.Б., Якир М.С. Название: Геометрия. 8 класс Формат: PDF Размер: 23,38 Мб Язык: Русский.
Скачать по прямой ссылке. Учебник предназначен для изучения геометрии в 8 классе общеобразовательных учреждений.www.psyoffice.ru
- ГДЗ решебник по геометрии 8 класс Мерзляк А.Г., Полонский…
«ГДЗ по геометрии 8 класс Дидактические материалы Мерзляк (Вентана-граф)» обеспечит восьмиклассникам всеобъемлющую помощь в учебном процессе по представленной технической дисциплине, с помощью верных ответов и подробно расписанных комментариев автора к решению каждого номера из учебника. Решебник позволит школьнику в кратчайшие сроки нейтрализовать имеющиеся проблемы с отставанием от школьной программы и заполнить накопившиеся пробелы в знаниях.
spishi.ltd
- ГДЗ Мерзляка | гдз (решебники) в свободном доступе к учебникам…
…материалам по математике, алгебре, геометрии Мерзляка А.Г., Полонского В.Б.
, Якира М.С. дляРешения заданий (ГДЗ) номер 1 и 2 из учебника Алгебра 8 класс Мерзляк Полонский.
в свободном доступе к учебникам, дидактическим материалам по математике, алгебре…
vk.com
- Геометрия 8 Контрольные Мерзляк | + ОТВЕТЫ
Геометрия 8 Контрольные Мерзляк — контрольные работы (цитаты) из пособия «Дидактические материалы 8 класс ФГОС» (Мерзляк Полонский Якир) с ответами.
8 класс ФГОС » (авт. А.Г. Мерзляк, В.Б. Полонский, М.С. Якир , изд-во «Вентана-Граф») для ознакомления, а также ОТВЕТЫ на них (в пособии нет ответов). Цитаты из указанного учебного пособия использованы на сайте в незначительных объемах, исключительно в учебных и информационных целях (пп. 1 п. 1 ст. 1274 ГК РФ): цитаты переработаны в удобный формат (каждая работа на 1-й странице)…
xn--b1agatflbfbtgq5jm.xn--p1ai
 net
net 8 класс» (авт. А.Г. Мерзляк, В.Б. Полонский, М.С. Якир) системы «Алгоритм успеха».
8 класс» (авт. А.Г. Мерзляк, В.Б. Полонский, М.С. Якир) системы «Алгоритм успеха».

 Г. Геометрия. Дидактические материалы. 8 класс.»
Г. Геометрия. Дидактические материалы. 8 класс.» Б. Полонский, М.С. Якир…
Б. Полонский, М.С. Якир… Г. Мерзляк, В.Б. Полонский
Г. Мерзляк, В.Б. Полонский
 Цитаты из методического пособия Е.В.Буцко.
Цитаты из методического пособия Е.В.Буцко. ..
.. Г, М52 Геометрия : 8 класс ; учебник для учащихся общеобразовательных учреждений / А.Г. Мерзляк, В.Б. Полонский, М.С. Якир.
Г, М52 Геометрия : 8 класс ; учебник для учащихся общеобразовательных учреждений / А.Г. Мерзляк, В.Б. Полонский, М.С. Якир. С. Якир , изд-во «Вентана-Граф»), а также РЕШЕНИЯ и ОТВЕТЫ (в пособии нет ответов). Цитаты из указанного учебного пособия использованы на сайте в незначительных объемах, исключительно в учебных и информационных целях (пп. 1 п. 1 ст. 1274 ГК РФ): цитаты переработаны в удобный формат…
С. Якир , изд-во «Вентана-Граф»), а также РЕШЕНИЯ и ОТВЕТЫ (в пособии нет ответов). Цитаты из указанного учебного пособия использованы на сайте в незначительных объемах, исключительно в учебных и информационных целях (пп. 1 п. 1 ст. 1274 ГК РФ): цитаты переработаны в удобный формат…
 Главное не злоупотреблять бессмысленным списыванием.
Главное не злоупотреблять бессмысленным списыванием. Г. Геометрия : дидактические материалы : 8 класс : пособие для учащихся общеобразовательных организаций / А.Г. Мерзляк, В.Б. Полонский, Е.М. Рабинович и др.
Г. Геометрия : дидактические материалы : 8 класс : пособие для учащихся общеобразовательных организаций / А.Г. Мерзляк, В.Б. Полонский, Е.М. Рабинович и др. Скачать по прямой ссылке. Учебник предназначен для изучения геометрии в 8 классе общеобразовательных учреждений.
Скачать по прямой ссылке. Учебник предназначен для изучения геометрии в 8 классе общеобразовательных учреждений. , Якира М.С. для
, Якира М.С. дляНа данной странице Вы можете найти лучшие результаты поиска для чтения, скачивания и покупки на интернет сайтах материалов, документов, бумажных и электронных книг и файлов похожих на материал «Геометрия, Дидактические материалы, 8 класс, Мерзляк А. Г., Полонский В.Б., Рабинович Е.М., Якир М.С., 2018»
Г., Полонский В.Б., Рабинович Е.М., Якир М.С., 2018»
Для формирования результатов поиска документов использован сервис Яндекс.XML.
Нашлось 33 млн ответов. Показаны первые 32 результата(ов).
Дата генерации страницы:
Онлайн решебники (гдз) по Геометрии 8 класса
- ГДЗ
- 8 класс
- Геометрия
Геометрия 8 класс дидактические материалы Б.
Г. ЗивАвторы: Б.Г. Зив В.М. Мейлер
Геометрия 7-9 класс Л.С. Атанасян
Авторы: Л.С. Атанасян В.Ф. Бутузов
Геометрия 7-9 класс А.В. Погорелов
Автор: А.В. Погорелов
Геометрия 8 класс В.В. Шлыков
Автор: В.В. Шлыков
Геометрия 8 класс рабочая тетрадь Л.
С. АтанасянАвторы: Л.С. Атанасян В.Ф. Бутузов
Геометрия 8 класс опорные конспекты Казаков В.В.
Автор: Казаков В.В.
Геометрия 8 класс Г.В. Апостолова
Автор: Г.В. Апостолова
Геометрия 8 класс Г.П. Бевз
Авторы: Г.П. Бевз В.Г. Бевз
Геометрия 8 класс А.
П. ЕршоваАвторы: А.П. Ершова В.В. Голобородько
 Г. Зив
Г. Зив С. Атанасян
С. Атанасян П. Ершова
П. ЕршоваГеометрия 8 класс М.І. Бурда
Авторы: М.І. Бурда Н.А. Тарасенкова
Геометрия 8 класс Мерзляк А.Г.
Авторы: Мерзляк А.Г. Полонський В.Б.
Геометрия 8 класс В.Б. Полонський
Авторы: В.Б. Полонський Ю.М. Рабінович
Геометрия 8 класс углубленное изучение Мерзляк А.
Г. Углубленный уровеньАвторы: Мерзляк А.Г. Полонський В.Б.
Геометрия 7-9 класс Погорєлов О.В.
Автор: Погорєлов О.В.
Геометрия 8 класс рабочая тетрадь Дудницын Ю.П.
Автор: Дудницын Ю.П.
Геометрия 8 класс А.Г. Мерзляк
Авторы: А.Г. Мерзляк В.Б. Полонский
Геометрия 8 класс дидактические материалы Мерзляк А.
Г.Авторы: Мерзляк А.Г. Полонский В.Б.
Геометрия 8 класс рабочая тетрадь Мерзляк А.Г.
Авторы: Мерзляк А.Г. Полонский В.Б.
 Г. Углубленный уровень
Г. Углубленный уровень Г.
Г.Геометрия 8 класс рабочая тетрадь Глазков Ю.А.
Авторы: Глазков Ю.А. Камаев П.М.
Геометрия 7-9 класс самостоятельные и контрольные работы Иченская М.А.
Автор: Иченская М.А.
Геометрия 8 класс контрольно-измерительные материалы Гаврилова Н.
Ф.Автор: Гаврилова Н.Ф.
Геометрия 8 класс Казаков В.В.
Автор: Казаков В.В.
Геометрия 8 класс Мерзляк А.Г. Углубленный уровень
Авторы: Мерзляк А.Г. Поляков В.М.
Геометрия 8 класс рабочая тетрадь Мищенко Т.М.
Автор: Мищенко Т.М.
Геометрия 8 класс рабочая тетрадь Универсальные учебные действия Глазков Ю.
А.Авторы: Глазков Ю.А. Егупова М.В.
Геометрия 8 класс тестовый контроль знаний Гальперина А.Р.
Автор: Гальперина А.Р.
Геометрия 8 класс контрольные работы Мельникова Н.Б.
Автор: Мельникова Н.Б.
 Ф.
Ф. А.
А.Геометрия 8 класс Шыныбеков А.Н.
Авторы: Шыныбеков А.Н. Шыныбеков Д.А.
Геометрия 8 класс Смирнов В.
А.Авторы: Смирнов В.А. Туяков Е.А.
Геометрия 8 класс Солтан Г.Н.
Авторы: Солтан Г.Н. Солтан А.Е.
Геометрия 8 класс тесты А. В. Фарков
Автор: А. В. Фарков
Геометрия 8 класс тесты Л.И. Звавич
Авторы: Л.И. Звавич Е.В. Потоскуев
Геометрия 8 класс контрольные измерительные материалы А.
Р. РязановскийАвторы: А. Р. Рязановский Д. Г. Мухин
 А.
А. Р. Рязановский
Р. Рязановский Для достижения высоких результатов по геометрии восьмикласснику вовсе не обязательно тратить все свободное время на ее изучение. Нанимать репетиторов или платить учителям в школе за дополнительные занятия после уроков тоже нет никакой необходимости. Простой способ повысить успеваемость – приобрести пособие с уже решенными заданиями. Кроме ответов на задания в такой книге есть и теоретический материал, изложенный простым и понятным для ученика языком, что поможет ему еще раз вспомнить объяснения учителя и разложить их в своей голове по полочкам. В пособии приведены как простые, так и сложные задания по геометрии. Есть даже так называемые «задания со звездочкой» — задачки повышенного уровня сложности. Просмотрев алгоритм их решения и разобравшись с порядком действий, ученик без труда сможет по аналогии решить такую задачку в классе во время контрольной работы. Все задачи разбиты на блоки, соответствующие темам и разделам учебника, так что найти нужный материал ученик сможет без проблем. Найти площадь, периметр, параметры окружности или любой другой фигуры ученик теперь сможет без особого труда, а если сложности все-таки возникнут, то ему нужно будет всего лишь заглянуть в пособие и еще раз вспомнить, как это делается.
Все задачи разбиты на блоки, соответствующие темам и разделам учебника, так что найти нужный материал ученик сможет без проблем. Найти площадь, периметр, параметры окружности или любой другой фигуры ученик теперь сможет без особого труда, а если сложности все-таки возникнут, то ему нужно будет всего лишь заглянуть в пособие и еще раз вспомнить, как это делается.
ГДЗ по геометрии для 8 класса Мерзляк А.Г.
Вопросы. Параграф. Решения
1 2 3 4 5 6 7 8 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 Решения»>
Параграф 1. Решения 1.1
1.2
1.3
1.4
1.5
1.6
1.7
1.8
1.9
1.10
1.11
1.12
1.13
1.14
1.15
1.16
1.17
1.18
1.19
1.20
1.21
1.22
1.23
1.24
1.25
1.26
1.
Решения»>
Параграф 1. Решения 1.1
1.2
1.3
1.4
1.5
1.6
1.7
1.8
1.9
1.10
1.11
1.12
1.13
1.14
1.15
1.16
1.17
1.18
1.19
1.20
1.21
1.22
1.23
1.24
1.25
1.26
1. 27
1.28
1.29
1.30
1.31
1.32
1.33
27
1.28
1.29
1.30
1.31
1.32
1.33Параграф 2. Решения
2.1 2.2 2.3 2.4 2.5 2.6 2.7 2.8 2.9 2.10 2.11 2.12 2.13 2.14 2.15 2.16 2.17 2.18 2.19 2. 20
2.21
2.22
2.23
2.24
2.25
2.26
2.27
2.28
2.29
2.30
2.31
2.32
2.33
2.34
2.35
2.36
2.37
2.38
2.39
2.40
2.41
2.42
2.43
2.44
2.45
2.46
2.47
2.48
2.
20
2.21
2.22
2.23
2.24
2.25
2.26
2.27
2.28
2.29
2.30
2.31
2.32
2.33
2.34
2.35
2.36
2.37
2.38
2.39
2.40
2.41
2.42
2.43
2.44
2.45
2.46
2.47
2.48
2. 49
49Параграф 3. Решения
3.1 3.2 3.3 3.4 3.5 3.6 3.7 3.8 3.9 3.10 3.11 3.12 3.13 3.14 3.15 3.16 3.17 3.18 3.19 3.20 3.21 3.22 3.23 3.24 3.25 3. 26
3.27
3.28
26
3.27
3.28Параграф 4. Решения
4.1 4.2Параграф 5. Решения
5.1 5.2 5.3 5.4 5.5 5.6 5.7 5.8 5.9 5.10 5.11 5.12 5.13 5.14 5.15 5.16 5.17 5. 18
5.19
5.20
5.21
5.22
5.23
5.24
5.25
5.26
5.27
5.28
5.29
5.30
5.31
5.32
5.33
5.34
5.35
5.36
5.37
5.38
5.39
5.40
5.41
5.42
5.43
5.44
5.45
5.46
5.
18
5.19
5.20
5.21
5.22
5.23
5.24
5.25
5.26
5.27
5.28
5.29
5.30
5.31
5.32
5.33
5.34
5.35
5.36
5.37
5.38
5.39
5.40
5.41
5.42
5.43
5.44
5.45
5.46
5. 47
5.48
5.49
5.50
5.51
5.52
5.53
5.54
5.55
5.56
5.57
5.58
47
5.48
5.49
5.50
5.51
5.52
5.53
5.54
5.55
5.56
5.57
5.58Параграф 6. Решения
6.1 6.2 6.3 6.4 6.5 6.6 6.7 6.8 6.9 6.10 6.11 6.12 6.13 6.14 6. 15
6.16
6.17
6.18
6.19
6.20
6.21
6.22
6.23
6.24
6.25
6.26
6.27
6.28
6.29
6.30
6.31
6.32
6.33
15
6.16
6.17
6.18
6.19
6.20
6.21
6.22
6.23
6.24
6.25
6.26
6.27
6.28
6.29
6.30
6.31
6.32
6.33Параграф 7. Решения
7.1 7.2 7.3 7.4 7.5 7.6 7.7 7. 8
7.9
7.10
7.11
7.12
7.13
7.14
7.15
7.16
7.17
7.18
7.19
7.20
7.21
7.22
7.23
7.24
7.25
7.26
7.27
7.28
7.29
7.30
7.31
7.32
7.33
7.34
7.35
7.36
7.
8
7.9
7.10
7.11
7.12
7.13
7.14
7.15
7.16
7.17
7.18
7.19
7.20
7.21
7.22
7.23
7.24
7.25
7.26
7.27
7.28
7.29
7.30
7.31
7.32
7.33
7.34
7.35
7.36
7. 37
7.38
7.39
7.40
7.41
7.42
7.43
7.44
37
7.38
7.39
7.40
7.41
7.42
7.43
7.44Параграф 8. Решения
8.1 8.2 8.3 8.4 8.5 8.6 8.7 8.8 8.9 8.10 8.11 8.12 8.13 8.14 8.15 8.16 8.17 8.18 8. 19
8.20
8.21
8.22
8.23
8.24
8.25
8.26
8.27
8.28
8.29
8.30
8.31
8.32
8.33
8.34
8.35
8.36
8.37
8.38
8.39
8.40
8.41
8.42
8.43
8.44
8.45
8.46
8.47
8.
19
8.20
8.21
8.22
8.23
8.24
8.25
8.26
8.27
8.28
8.29
8.30
8.31
8.32
8.33
8.34
8.35
8.36
8.37
8.38
8.39
8.40
8.41
8.42
8.43
8.44
8.45
8.46
8.47
8. 48
8.49
8.50
8.51
8.52
8.53
8.54
8.55
8.56
8.57
8.58
8.59
8.60
48
8.49
8.50
8.51
8.52
8.53
8.54
8.55
8.56
8.57
8.58
8.59
8.60Параграф 9. Решения
9.1 9.2 9.3 9.4 9.5 9.6 9.7 9.8 9.9 9.10 9.11 9.12 9.13 9. 14
9.15
9.16
9.17
9.18
9.19
9.20
9.21
9.22
9.23
9.24
9.25
9.26
9.27
9.28
9.29
9.30
9.31
9.32
14
9.15
9.16
9.17
9.18
9.19
9.20
9.21
9.22
9.23
9.24
9.25
9.26
9.27
9.28
9.29
9.30
9.31
9.32Параграф 10. Решения
10.1 10.2 10.3 10.4 10.5 10.6 10.7 10. 8
10.9
10.10
10.11
10.12
10.13
10.14
10.15
10.16
10.17
10.18
10.19
10.20
10.21
10.22
10.23
10.24
10.25
10.26
10.27
10.28
10.29
10.30
10.31
10.32
10.33
10.34
10.35
10.
8
10.9
10.10
10.11
10.12
10.13
10.14
10.15
10.16
10.17
10.18
10.19
10.20
10.21
10.22
10.23
10.24
10.25
10.26
10.27
10.28
10.29
10.30
10.31
10.32
10.33
10.34
10.35
10. 36
10.37
10.38
10.39
36
10.37
10.38
10.39Параграф 11. Решения
11.1 11.2 11.3 11.4 11.5 11.6 11.7 11.8 11.9 11.10 11.11 11.12 11.13 11.14 11.15 11.16 11.17 11.18 11.19 11.20 11.21 Решения»>
Параграф 12. Решения 12.1
12.2
12.3
12.4
12.5
12.6
12.7
12.8
12.9
12.10
12.11
12.12
12.13
12.14
12.15
12.16
12.17
12.18
12.19
12.20
12.21
12.22
12.23
12.24
12.25
12.
Решения»>
Параграф 12. Решения 12.1
12.2
12.3
12.4
12.5
12.6
12.7
12.8
12.9
12.10
12.11
12.12
12.13
12.14
12.15
12.16
12.17
12.18
12.19
12.20
12.21
12.22
12.23
12.24
12.25
12. 26
12.27
12.28
12.29
12.30
12.31
26
12.27
12.28
12.29
12.30
12.31Параграф 13. Решения
13.1 13.2 13.3 13.4 13.5 13.6 13.7 13.8 13.9 13.10 13.11 13.12 13.13 13.14 13.15 13.16 13.17 13.18 13.19 13. 20
13.21
13.22
13.23
13.24
20
13.21
13.22
13.23
13.24Параграф 14. Решения
14.1 14.2 14.3 14.4 14.5 14.6 14.7 14.8 14.9 14.10 14.11 14.12 14.13 14.14 14.15 14.16 14.17 14.18 14.19 14.20 14. 21
14.22
21
14.22Параграф 15. Решения
15.1 15.2 15.3 15.4 15.5 15.6 15.7 15.8 15.9 15.10 15.11 15.12 15.13 15.14 15.15 15.16 15.17 15.18 15.19 15.20 15.21 15.22 15.23 15. 24
15.25
15.26
15.27
15.28
15.29
15.30
15.31
15.32
15.33
15.34
15.35
15.36
15.37
15.38
15.39
15.40
15.41
15.42
15.43
15.44
15.45
15.46
15.47
15.48
15.49
15.50
15.51
15.
24
15.25
15.26
15.27
15.28
15.29
15.30
15.31
15.32
15.33
15.34
15.35
15.36
15.37
15.38
15.39
15.40
15.41
15.42
15.43
15.44
15.45
15.46
15.47
15.48
15.49
15.50
15.51
15. 52
15.53
52
15.53Параграф 16. Решения
16.1 16.2 16.3 16.4 16.5 16.6 16.7 16.8 16.9 16.10 16.11 16.12 16.13 16.14 16.15 16.16 16.17 16.18 16.19Параграф 17. Решения
17.1 17. 2
17.3
17.4
17.5
17.6
17.7
17.8
17.9
17.10
17.11
17.12
17.13
17.14
17.15
2
17.3
17.4
17.5
17.6
17.7
17.8
17.9
17.10
17.11
17.12
17.13
17.14
17.15Параграф 18. Решения
18.1 18.2 18.3 18.4 18.5 18.6 18.7 18.8 18.9 18.10 18.11 18. 12
18.13
18.14
18.15
18.16
18.17
18.18
18.19
18.20
18.21
18.22
18.23
18.24
18.25
12
18.13
18.14
18.15
18.16
18.17
18.18
18.19
18.20
18.21
18.22
18.23
18.24
18.25Параграф 19. Решения
19.1 19.2 19.3 19.4 19.5 19.6 19.7 19.8 19.9 19.10 19.11 19. 12
19.13
19.14
19.15
19.16
19.17
19.18
19.19
19.20
19.21
19.22
19.23
19.24
19.25
12
19.13
19.14
19.15
19.16
19.17
19.18
19.19
19.20
19.21
19.22
19.23
19.24
19.25Параграф 20. Решения
20.1 20.2 20.3 20.4 20.5 20.6 20.7 20.8 20.9 20.10 20.11 20. 12
20.13
20.14
20.15
20.16
20.17
20.18
20.19
20.20
20.21
20.22
20.23
20.24
20.25
20.26
20.27
20.28
20.29
20.30
20.31
20.32
20.33
20.34
20.35
20.36
20.37
20.38
20.39
20.
12
20.13
20.14
20.15
20.16
20.17
20.18
20.19
20.20
20.21
20.22
20.23
20.24
20.25
20.26
20.27
20.28
20.29
20.30
20.31
20.32
20.33
20.34
20.35
20.36
20.37
20.38
20.39
20. 40
20.41
20.42
20.43
20.44
20.45
20.46
20.47
20.48
40
20.41
20.42
20.43
20.44
20.45
20.46
20.47
20.48Параграф 21. Решения
21.1 21.2 21.3 21.4 21.5 21.6 21.7 21.8 21.9 21.10 21.11 21.12 21.13 21.14 21.15 21.16 21. 17
21.18
21.19
21.20
21.21
21.22
21.23
21.24
21.25
21.26
21.27
21.28
21.29
17
21.18
21.19
21.20
21.21
21.22
21.23
21.24
21.25
21.26
21.27
21.28
21.29Параграф 22. Решения
22.1 22.2 22.3 22.4 22.5 22.6 22.7 22.8 22.9 22.10 22.11 22.12 22. 13
22.14
22.15
22.16
22.17
22.18
22.19
22.20
22.21
22.22
22.23
22.24
22.25
22.26
22.27
22.28
13
22.14
22.15
22.16
22.17
22.18
22.19
22.20
22.21
22.22
22.23
22.24
22.25
22.26
22.27
22.28Параграф 23. Решения
23.1 23.2 23.3 23.4 23.5 23.6 23.7 23.8 23.9 23. 10
23.11
23.12
23.13
23.14
10
23.11
23.12
23.13
23.14Параграф 24. Решения
24.1 24.2 24.3 24.4 24.5 24.6 24.7 24.8 24.9 24.10 24.11 24.12 24.13 24.14 24.15Параграф 25. Решения
25.1 25.2 25. 3
25.4
25.5
25.6
25.7
25.8
25.9
25.10
25.11
25.12
25.13
25.14
25.15
25.16
25.17
25.18
25.19
25.20
25.21
25.22
25.23
25.24
25.25
25.26
25.27
25.28
25.29
25.30
25.
3
25.4
25.5
25.6
25.7
25.8
25.9
25.10
25.11
25.12
25.13
25.14
25.15
25.16
25.17
25.18
25.19
25.20
25.21
25.22
25.23
25.24
25.25
25.26
25.27
25.28
25.29
25.30
25. 31
25.32
25.33
25.34
25.35
25.36
25.37
25.38
25.39
25.40
25.41
25.42
25.43
25.44
25.45
25.46
25.47
25.48
25.49
25.50
25.51
25.52
25.53
25.54
25.55
25.56
25.57
25.58
31
25.32
25.33
25.34
25.35
25.36
25.37
25.38
25.39
25.40
25.41
25.42
25.43
25.44
25.45
25.46
25.47
25.48
25.49
25.50
25.51
25.52
25.53
25.54
25.55
25.56
25.57
25.58 Решения»>
Параграф 26. Решения 26.1
26.2
26.3
26.4
26.5
26.6
26.7
26.8
26.9
26.10
26.11
26.12
26.13
26.14
26.15
26.16
26.17
26.18
26.19
26.20
26.21
26.22
26.23
26.24
26.25
26.
Решения»>
Параграф 26. Решения 26.1
26.2
26.3
26.4
26.5
26.6
26.7
26.8
26.9
26.10
26.11
26.12
26.13
26.14
26.15
26.16
26.17
26.18
26.19
26.20
26.21
26.22
26.23
26.24
26.25
26. 26
26.27
26.28
26.29
26.30
26.31
26.32
26.33
26.34
26.35
26.36
26
26.27
26.28
26.29
26.30
26.31
26.32
26.33
26.34
26.35
26.36по геометрии за 8 класс Мерзляк, Полонский, Якир. Учебник ФГОС Вентана-Граф
по географии 8 класс мерзляк учебник номер 492
Геометрия – очень увлекательный предмет. Учащиеся восьмого класса должны обладать по нему некоторым багажом знаний. Впрочем, не будем скрывать, что не каждому подростку премудрости данной науки даются легко. Предмет потребует длительных усилий и прилежания.
Авторы-разработчики Мерзляк и другие разработали прекрасный учебно-методический комплекс. Подготовкой его к печати, дизайном обложки и иллюстрированием занимается «Просвещение». На обложке книги изображены красочные витиеватые фигуры, которые сразу же приковывают взгляд и мотивируют заниматься.
Решебник призван помочь школьникам, которые хотели бы лучше понимать предмет. Подобранные задачи будут полезны в закреплении и повторении знаний, понимании способов доказательства теорем. Они систематически отрабатывают образное мышление. При желании по этим можно подготовиться и к ОГЭ по математике.
Почему многие учащиеся выбирают такого помощника по геометрии?
В книгу были включены все подробные алгоритмы, необходимые для верного решения. Любой школьник разберется в тонкостях поиска углов и длин сторон треугольников, параллелограммов, трапеций и квадратов. Результатом станет уверенность в себе, высокие баллы на контрольных и проверочных работах. Преимущества использования Решебника по геометрии для 8 класса (авторы: А. Г. Мерзляк, В. Б. Полонский, М. С. Якир) следующие:
- легкий поиск по специальному указателю; верные примеры оформления графических задач; информативные комментарии; доступность материалов с любого подключенного в Интернет телефона.

Положительный результат придет уже в скором времени. Не стоит откладывать учебу на неопределенные сроки. Лучше быстрее взяться за подготовку и ощутить все положительные черты занятий по решебнику уже сейчас. Никакая теорема Пифагора или Герона не сможет остановить мотивированного школьника на пути к успеху и отличным отметкам в дневнике.
Сборник по геометрии под авторством А. Г. Мерзляка пригодится для улучшения успеваемости по математике, повторения материала перед проверочными испытаниями, пробниками ОГЭ. Частные репетиторы могут использовать приведенные упражнения с ответами в своих методиках.
Похожие
Дидактические материалы геометрия 8 класс Мерзляк А. Г.
Математические диктанты, Контрольные работы (Методическое пособие) геометрия 8 класс Буцко Е. В.
Геометрия очень увлекательный предмет.
Megaresheba. ru
21.04.2017 7:15:10
2017-04-21 07:15:10
Источники:
Https://megaresheba. ru/gdz/geometriya/8-klass/merzlyak
8 клас 2021 рік (відповіді до підручників) » /> » /> . keyword { color: red; }
keyword { color: red; }
по географии 8 класс мерзляк учебник номер 492
Навчання у 8 класі передбачає вивчення таких, не зовсім улюблених багатьма школярами, предметів як алгебра, геометрія, фізика, а також хімія. Звичайно, є такі учні, яким осягнення тонкощів таких наук дається просто. Правда, їх незначне число. А що робити всім іншим? Адже більшість таких предметів необхідно надалі здавати як при державній атестації, так і при вступі для здобуття вищої освіти.
Варто відзначити, що найкраще в подібному випадку не упускати момент і виконувати вправи, активно застосовуючи при цьому готові домашні завдання за 8 клас. Звичайно, багато учнів, яким не вдається виконати завдання з того чи іншого предмету просто закидають вивчення подібної дисципліни надалі. Особливо часто таке може спостерігатися в тому випадку, коли викладач часто виставляє негативні оцінки. Особливо, за неправильно виконану домашню роботу.
Багатьом батькам і навіть самим школяра здається, що розірвати дане коло практично неможливо. Але, насправді це далеко не так. Тим більше, що з появою різноманітних збірників, де розміщені повністю розв’язання з англійської мови, біології, географії, а також української літератури можна досить успішно вирішити цю проблему. Найголовніше стежити за тим, чи представлений в розв’язнику матеріал саме того підручника, по якому ви займаєтеся.
Але, насправді це далеко не так. Тим більше, що з появою різноманітних збірників, де розміщені повністю розв’язання з англійської мови, біології, географії, а також української літератури можна досить успішно вирішити цю проблему. Найголовніше стежити за тим, чи представлений в розв’язнику матеріал саме того підручника, по якому ви займаєтеся.
Не секрет, що сучасні шкільні вчителі найчастіше ставлять дуже великий обсяг завдань. Причому, далеко не кожен учень може саме на уроці засвоїти весь той матеріал, який необхідний для якісного виконання кожної задачі. Тому, найбільш оптимальним способом і надолужити все упущене і при цьому зробити всі завдання — . Багато хто розглядає такий варіант лише як спосіб вирішення задач і вправ, заданих додому. Але це не зовсім так.
У восьмому класі багато викладачів дають вельми складні завдання, які без хорошої попередньої підготовки не вирішити. А в тому випадку, коли ви маєте в своєму розпорядженні відповіді з того чи іншого предмету, то цілком можливо потренуватися у здійсненні успішного вирішення навіть найскладнішого доручення. Головне, мати відповідне бажання.
Головне, мати відповідне бажання.
Copyright @ 2014-2022 до підручників і робочих зошитів, готові домашні завдання, решебник, розв’язані задачі та вправи для 1-11 класів. | Контакти
Адже більшість таких предметів необхідно надалі здавати як при державній атестації, так і при вступі для здобуття вищої освіти.
Gdzonline. net
27.01.2018 18:12:23
2018-01-27 18:12:23
Источники:
Https://gdzonline. net/8_klas/
Номер (задание) 492 — по геометрии 8 класс Мерзляк, Полонский, Якир » /> » /> .keyword { color: red; }
по географии 8 класс мерзляк учебник номер 492
Подробный решебник () по Геометрии за 8 (восьмой) класс — готовый ответ номер — 492. Авторы учебника: Мерзляк, Полонский, Якир. Издательство: Вентана-граф 2016.
Похожие
Дидактические материалы геометрия 8 класс Мерзляк А. Г.
Математические диктанты, Контрольные работы (Методическое пособие) геометрия 8 класс Буцко Е. В.
В.
Рабочая тетрадь геометрия 8 класс Мерзляк А. Г.
учебник геометрия 8 класс Мерзляк А. Г. углубленный уровень
492. Отрезки АВ и CD пересекаются в точке О (рис. 165), АО = 24 см, ВО = 16 см, СО = 15 см, OD = 10 см, ∠ACO = 72°. Найдите ∠BDO.
Дидактические материалы геометрия 8 класс Мерзляк А.
Megaresheba. ru
05.11.2018 5:09:35
2018-11-05 05:09:35
Источники:
Https://megaresheba. ru/gdz/geometriya/8-klass/merzlyak/492-nomer
Тесты по геометрии для 8 класса онлайн
Окружность
-
ВПР математика 8 класс
04.05.2020 4155 0
Назначение ВПР по математике — оценить уровень общеобразовательной подготовки обучающихся 8 класса. В работе 19 заданий.
14 заданий курса алгебре и 5 заданий курса геометрии. Максимальное количество баллов 25. -
Прямоугольник. 8 класс
07.10.2021 1718
Тест для контроля знаний учащихся по теме «Прямоугольник». Геометрия 8 класс.
-
Четырехугольники и их свойства. 8 класс.
06.11.2020 2146
Тест предназначен для проверки теоретических знаний по теме » Четырёхугольники и их свойства.» 8 класс.Тест ориентирован на учебник «Геометрия 7-9 классы » Л.С.Атанасян и др. В тесте 44 вопроса. Для ответа случайным образом выбирается 20 вопросов.
-
ВПР-2020.
8 класс. Математика. Тест 1
07.02.2020 6064 0
Тест 1 — подготовка к Всероссийской проверочной работе по математике. 8 класс.
-
Итоговый тест 8 класс геометрия ТЕОРИЯ
08.05.2021 3383 0
Тест содержит 30 заданий. Задания в тест выбираются случайным образом из общей базы заданий. Критерии: «3» 50-69%, «4» 70-90%, «5» 91-100%. Оценка выставляется сразу после прохождения теста.
-
8 класс. Геометрия. Центральные и вписанные углы.
25.07.2017 19683
Тест предназначен для учащихся 8 классов при отработке навыков решения задач по теме «Центральные и вписанные углы».
-
Векторы на плоскости Основные понятия.
31.10.2021 894 0
Тест содержит вопросы на основные определения, связанные с векторами, а также на действия с векторами на плоскости.
-
8 класс Геометрия
15.03.2019 29282
Данный тест предназначен для определения знания предмета «Геометрия» за курс 8 класса.
-
ГЕОМЕТРИЯ выбор верных утверждений 8 класс
12.04.2021 4509 0
Тест содержит 30 заданий.
Задания в тест выбираются случайным образом из общей базы заданий. Критерии: «3» 50-69%, «4» 70-90%, «5» 91-100%. Оценка выставляется сразу после прохождения теста. Время ОГРАНИЧЕНО 20 минут. -
Признаки подобия треугольников. 8 класс
07.02.2022 1558
Тест «Признаки подобия треугольников» состоит из трех вопросов (задач), выпадающих случайным образом из общего списка.
-
Входной тест Геометрия 8
23.07.2020 183 0
Входной тест по геометрии, нацеленный на проверку остаточных знаний по геометрии в начале учебного года, проверяет программу по геометрии 7 класса.
-
8 класс. Геометрия. Соотношения в прямоугольном треугольнике
04.04.2015 8537
Тест предназначен для учащихся 8 классов при отработке навыков решения задач по теме «Соотношения в прямоугольном треугольнике».
 14 заданий курса алгебре и 5 заданий курса геометрии. Максимальное количество баллов 25.
14 заданий курса алгебре и 5 заданий курса геометрии. Максимальное количество баллов 25. 8 класс. Математика. Тест 1
8 класс. Математика. Тест 1

 Задания в тест выбираются случайным образом из общей базы заданий. Критерии: «3» 50-69%, «4» 70-90%, «5» 91-100%. Оценка выставляется сразу после прохождения теста. Время ОГРАНИЧЕНО 20 минут.
Задания в тест выбираются случайным образом из общей базы заданий. Критерии: «3» 50-69%, «4» 70-90%, «5» 91-100%. Оценка выставляется сразу после прохождения теста. Время ОГРАНИЧЕНО 20 минут.
-
8 класс Глава 5 Четырёхугольники ТЕОРИЯ
09.05.2021 2523 0
Тест содержит 20 заданий. Задания в тест выбираются случайным образом из общей базы заданий. Критерии: «3» 50-69%, «4» 70-90%, «5» 91-100%. Оценка выставляется сразу после прохождения теста.
-
Тест № 1.
Тема «Окружность»
10.09.2022 4 0
Тест представляет собой теоретические вопросы по теме: «Окружность». Тест предназначен для определения предметных результатов учащихся 8 класса по теме.
-
Определение подобных треугольников. 8 класс
20.01.2020 8705 0
Самостоятельная работа в форме теста по теме: «Определение подобных треугольников»
-
Центральные и вписанные углы
29.04.2020 5679 0
Тест позволяет закрепить тему «Центральные и вписанные углы».
Тест содержит две задачи из кзаменационных материалов ОГЭ. -
Итоговая контрольная работа по математике за курс 8 класса. 8Б класс
08.05.2020 275 0
Итоговая работа соответствует матералам 8 класса, изучаемым по учебникам Алгебра 8 клаас (Колягин) и Геометрия 8 класс (Ататнасян). Работа содержит 10 заданий (6 по алгебре и 4 по геометрии). На выполнение работы отводится 1час 30 минут. Количество прохождений: 1.
-
Площади четырёхугольников. 8 класс.
07.11.2020 713 0
Тест предназначен для проверки теоретических знаний по теме » Площади четырёхугольников», 8 класс.
Тест ориентирован на учебник «Геометрия 7-9 классы » Л.С.Атанасян и др. В тесте 27 вопросов. Для ответа случайным образом выбирается 15 вопросов. Может быть использован для подготовки к ОГЭ по математике. -
ВПР-2020. 8 класс. Математика. Тест 2
13.02.2020 384 0
Тест 2 — подготовка к Всероссийской проверочной работе по математике. 8 класс.
-
Итоговый тест по геометрии, тема: «Решение треугольника, подобные треугольники»
27.04.2020 282 0
Проверочный тест к теме «Решение треугольников и подобные треугольники»
-
ОГЭ 8 класс математика
29.
04.2020
590
0
Вариант ОГЭ математика 2020 год. Содержит 25 демонстрационных вариантов из сборника подготовки к ОГЭ, адаптированных к программе 8 класса.
-
Площадь многоугольника.
20.02.2021 38 0
Проверочная работа по теме «Площадь многоугольника» предназначена для проверки знаний по указанной теме. Работа составлена на основе прототипов заданий ОГЭ по математике.
-
Формула Пика. Задачи на клетчатой бумаге.
24.11.2021 77 0
Данный тест будет полезен педагогу для проверки знаний по теме «Площади многоугольников» и ребятам для закрепления материала.
-
Тест по теме «Соотношения между сторонами и углами прямоугольного треугольника»
02.02.2022 1300 0
Данный тест проверяет знания по теме «Соотношения между сторонами и углами прямоугольного треугольника», геометрия, 8 класс
 Тема «Окружность»
Тема «Окружность»
 Тест содержит две задачи из кзаменационных материалов ОГЭ.
Тест содержит две задачи из кзаменационных материалов ОГЭ. Тест ориентирован на учебник «Геометрия 7-9 классы » Л.С.Атанасян и др. В тесте 27 вопросов. Для ответа случайным образом выбирается 15 вопросов. Может быть использован для подготовки к ОГЭ по математике.
Тест ориентирован на учебник «Геометрия 7-9 классы » Л.С.Атанасян и др. В тесте 27 вопросов. Для ответа случайным образом выбирается 15 вопросов. Может быть использован для подготовки к ОГЭ по математике. 04.2020
590
0
04.2020
590
0

-
Практические задачи на тему: «Подобие треугольников».
03.02.2022 620 0
Тестовые задания, проверяющие умение применять подобие треугольников для решения практических задач.
-
Викторина о тригонометрии
21.
02.2022
390
0
Викторина о разделе математики, изучающем функции угла: синус, косинус и т.п. В рамках школьной программы.
-
Площадь. 8 класс
14.12.2019 12273
Тест предназначен для повторения темы «Площадь». Базовый уровень сложности. 5 заданий. Время выполнения 15 минут.
-
Тест по заданию №17 (ОГЭ-2020 по математике) по теме: Решение геометрических задач по теме «Окружность и круг».
16.02.2020 863 0
Тест содержит десять тестовых заданий по основным прототипам задания №17 (ОГЭ 2020).
К каждому номеру ответом является число (без единиц измерения). -
Вписанные и центральные углы. 8 класс. урок 2. Повторение.
13.04.2020 140 0
Тетс составлен к теме «Вписанные и описанные углы» по учебнику Л.С.Атаносяна геометрия 8 класс.
-
свойство хорд и углов, образованных касательными и хордами
14.04.2020 221 0
Тест предназначен ддля первичного закрепления на уроке по теме «Свойство хорд окружности». Все задачи решаются устно с последующей самопроверкой
-
СООТНОШЕНИЯ В ПРЯМОУГОЛЬНОМ ТРЕУГОЛЬНИКЕ.
ТЕОРЕМА ПИФАГОРА
16.04.2020 3886 0
Тест по темам: «Соотношения в прямоугольном треугольника. Теорема Пифагора» для обучающихся в 8 классе.
-
8 класс. Центральные и вписанные углы
20.04.2020 556 0
Для успешного прохождения теста повторите, что такое центральные и вписанные углы, свойства биссектрисы угла. Время прохождения теста — 30 минут. Удачи!
-
Геометрия 8 класс. Окружность.
22.04.2020 217
Тест по теме: «Окружность».
Теоретические вопросы по данной теме. -
МД «Свойство биссектрисы угла»
23.04.2020 739 0
Решение задач по готовым чертежам по теме «Биссектриса угла» геометрия 8 класс по учебнику Атоносяна. Проводится в начале урока «Серединный перпендикуляр»
-
Вариант 2 Свойства биссектрисы угла и серединного перпендикуляра к отрезку
24.04.2020 643 0
Тест ориентирован на учебник Л.С. Атанасяна и др. «Геометрия. 7 — 9 классы». Направлен на первичный контроль текущего усвоения материала. Задания проверяют понимание новой терминологии, распознавание видов фигур и их свойств.
-
Площадь многоугольника.
24.04.2020 184 0
Тест предназначен для проверки знаний по геометрии по теме «Площадь».
 02.2022
390
0
02.2022
390
0
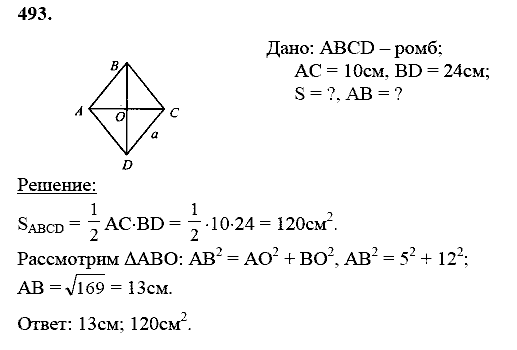 К каждому номеру ответом является число (без единиц измерения).
К каждому номеру ответом является число (без единиц измерения). ТЕОРЕМА ПИФАГОРА
ТЕОРЕМА ПИФАГОРА
 Теоретические вопросы по данной теме.
Теоретические вопросы по данной теме.
-
Центральный и вписанный углы
11.05.2020 23 0
Тест предназначен для самостоятельной работы с учебником. Задания составлены так, чтобы каждый участник мог в должной мере оценить знания по предмету, причем участвовать можно как группой, так и индивидуально.
-
Промежуточная аттестация
11.
05.2020
178
0
Данный тест предназначен для определения знания предмета «Геометрия» за курс 8 класса.
-
окружность_ геометрия_8класс
21.05.2020 539 0
Геометрия 8 класс. Тема Окружность. Учебник Атанасяна Л.С. и др. Итоговый тест
-
Центральные и вписанные углы
24.11.2020 111 0
Тест составлен на основе учебника по геометрии за 8 класс авторы Мерзляк А.Г., Полонский В. Б., Якир М. С. Параграф девять. Глава один
-
Контрольная работа за І семестр по геометрии 8 класс
11.
12.2020
14
0
Контрольная работа за I семестр по геометрии состоит из 10 заданий. с 1 по 6 — тестовые задания (от 1 до 3 правильных), с 7 по 8 — установить соответствие, 9 — расположить в правильной последовательности, 10 — указать только ответ (число).
-
Неравенство треугольника
17.12.2020 61 0
Тест предназначен для учащихся средней школы для проверки уровня знаний по теме «Неравенство треугольника».
-
Пропорциональные отрезки. Подобные треугольники.
07.02.2021 61 0
Тест составлен по вопросам к учебнику А.
Г. Мерзляк, В.Б Полонский. В тесте содержатся вопросы о свойстве биссектрисы треугольника, свойстве касательной и секущей, свойстве отрезков пересекающихся хорд. -
Углы, образованные хордами, секущими и касательными
14.03.2021 305 0
Тренировочный тест предназначен для учащихся 8-ых классов при отработке навыков решения задач по теме «Углы, образованные хордами, секущими и касательными».
-
Окружность (итоговый тест)
14.03.2021 1872 0
Тренировочный тест предназначен для учащихся 8-ых классов при отработке навыков решения задач по теме «Окружность».
-
8 класс Глава 8 Окружность ТЕОРИЯ
08.05.2021 773 0
Тест содержит 20 заданий. Задания в тест выбираются случайным образом из общей базы заданий. Критерии: «3» 50-69%, «4» 70-90%, «5» 91-100%. Оценка выставляется сразу после прохождения теста.
-
8 класс Глава 6 Площадь ТЕОРИЯ
09.05.2021 249 0
Тест содержит 20 заданий. Критерии: «3» 50-69%, «4» 70-90%, «5» 91-100%. Оценка выставляется сразу после прохождения теста.
-
8 класс Глава 7 Подобные треугольники ТЕОРИЯ
09.
05.2021
915
0
Тест содержит 20 заданий. Задания в тест выбираются случайным образом из общей базы заданий. Критерии: «3» 50-69%, «4» 70-90%, «5» 91-100%. Оценка выставляется сразу после прохождения теста.
 05.2020
178
0
05.2020
178
0
 12.2020
14
0
12.2020
14
0
 Г. Мерзляк, В.Б Полонский. В тесте содержатся вопросы о свойстве биссектрисы треугольника, свойстве касательной и секущей, свойстве отрезков пересекающихся хорд.
Г. Мерзляк, В.Б Полонский. В тесте содержатся вопросы о свойстве биссектрисы треугольника, свойстве касательной и секущей, свойстве отрезков пересекающихся хорд.
 05.2021
915
0
05.2021
915
0
-
Итоговый тест 8 класс геометрия ЗАДАЧИ
09.05.2021 2098 0
Тест содержит 20 заданий. Критерии: «3» 50-69%, «4» 70-90%, «5» 91-100%. Оценка выставляется сразу после прохождения теста.
-
геометрия к ОГЭ
09.08.2021 119 0
Данный тест составлен на основе вопросов ОГЭ по теме «Углы» по курсу геометрии 8 класса.
Он поможет проверит основные знания. -
Теорема Пифагора
10.11.2021 113 0
Тест по геометрии по теме «Теорема Пифагора» 8 класс по учебнику Погорелова
-
теорема Пифагора
10.11.2021 250 0
Тест по геометрии 8 класс по учебнику Погорелов по закреплению теорема Пифагора
-
Теорема Пифагора
10.11.2021 1269
тест по геометрии 8 класс , учебник Погорелов , по теме : Теорема Пифагора
-
Теорема Пифагора (домашнее задание №1)
22.
11.2021
364
0
Домашнее задание в тестовой форме. В тесте 5 вопросов. Система оценивания: «5» — 5 верно выполненных заданий (95%) «4» — 4 верно выполненных заданий (75%) «3» — 3 верно выполненных заданий (55%) «2» — 2 и менее верно выполненных заданий (меньше 55%)
-
Тест по теме «Четырехугольники» 8 класс
25.11.2021 66 0
Тест предназначен для учащихся 8 класса для проверки и закрепления знаний по теме «Четырехугольники»
-
Теорема Пифагора
04.12.2021 1153 0
Тест содержит 7 вопросов по геометрии,тема теорема Пифагора, предназначен для учеников 8 класса.
В тесте 2 задания с выбором ответа и 5 заданий с вводом числового ответа. Тест содержит простые задачи на умение находить гипотенузу и катет прямоугольного треугольника,по известным 2 сторонам. В нем есть более сложные задачи с использованием теоремы Пифагора и других формул и теорем геометрии, например, формул площадей и периметра геометрических фигур. В заданиях кроме прямоугольного треугольника надо знать и уметь применять свойства равнобедренного треугольника, прямоугольника, ромба. Вычислять площадь прямоугольника, прямоугольного треугольника, ромба. Уметь вычислять площадь ромба через диагонали. -
Тест № 2 по теме «Окружность»
10.09.2022 4 0
Тест содержит пятнадцать практических заданий по теме «Окружность» и предназначен для определения предметных результатов учащихся 8 класса.
 Он поможет проверит основные знания.
Он поможет проверит основные знания. 11.2021
364
0
11.2021
364
0
 В тесте 2 задания с выбором ответа и 5 заданий с вводом числового ответа. Тест содержит простые задачи на умение находить гипотенузу и катет прямоугольного треугольника,по известным 2 сторонам. В нем есть более сложные задачи с использованием теоремы Пифагора и других формул и теорем геометрии, например, формул площадей и периметра геометрических фигур. В заданиях кроме прямоугольного треугольника надо знать и уметь применять свойства равнобедренного треугольника, прямоугольника, ромба. Вычислять площадь прямоугольника, прямоугольного треугольника, ромба. Уметь вычислять площадь ромба через диагонали.
В тесте 2 задания с выбором ответа и 5 заданий с вводом числового ответа. Тест содержит простые задачи на умение находить гипотенузу и катет прямоугольного треугольника,по известным 2 сторонам. В нем есть более сложные задачи с использованием теоремы Пифагора и других формул и теорем геометрии, например, формул площадей и периметра геометрических фигур. В заданиях кроме прямоугольного треугольника надо знать и уметь применять свойства равнобедренного треугольника, прямоугольника, ромба. Вычислять площадь прямоугольника, прямоугольного треугольника, ромба. Уметь вычислять площадь ромба через диагонали.
Проводимость и ионная селективность мезоскопических белковых нанопор, зондированных с помощью сканирующего мутагенеза цистеина
1. Hille, B. 1992. Ионные каналы возбудимых мембран. Sinauer Associates, Сандерленд, Массачусетс.
2. Красильников О.В., Мерзляк П.Г., Сабиров Р.З., Ташмухамедов Б.А. 1990. Память — это свойство пула ионных каналов — ионных каналов, образованных Staphylococcus aureus α -токсином. Ген. физиол. Биофиз. 9: 569–575. [PubMed] [Академия Google]
3. Безруков С.М., Касьянович Дж.Дж. 1993. Текущий шум раскрывает кинетику протонирования и количество ионизируемых участков в открытом ионном канале белка. физ. Преподобный Летт. 70:2352–2355. [PubMed] [Google Scholar]
4. Kasianowicz, J.J., and S.M. Bezrukov. 1995. Динамика протонирования ионного канала α -токсина по данным спектрального анализа рН-зависимых флуктуаций тока. Биофиз. Дж. 69:94–105. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. Kasianowicz, JJ, E. Brandin, D. Branton, and D.W. Deamer. 1996. Характеристика индивидуальных полимуклеотидных молекул с помощью мембранного канала. проц. Натл. акад. Наука США. 93:13770–13773. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Kasianowicz, JJ, E. Brandin, D. Branton, and D.W. Deamer. 1996. Характеристика индивидуальных полимуклеотидных молекул с помощью мембранного канала. проц. Натл. акад. Наука США. 93:13770–13773. [Бесплатная статья PMC] [PubMed] [Google Scholar]
6. Корнелл, Б. А., В. Л. Б. Браах-Максвитис, Л. Г. Кинг, П. Д. Дж. Осман, Б. Рагузе, Л. Вечорек и Р. Дж. Пейс. 1997. Биосенсор, использующий переключатели ионных каналов. Природа. 387: 580–583. [PubMed] [Google Scholar]
7. Браха О., Б. Уокер, С. Чели, Дж. Дж. Касьянович, Л. З. Сонг, Дж. Э. Гуо и Х. Бейли. 1997. Разработаны белковые поры как компоненты биосенсоров. хим. биол. 4: 497–505. [PubMed] [Google Scholar]
8. Kasianowicz, JJ, D.L. Burden, LC Han, S. Cheley, and H. Bayley. 1999. Генетически сконструированные сайты связывания ионов металлов снаружи трансмембранного канала β -ствола. Биофиз. Дж. 76:837–845. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Kasianowicz, JJ, S.E. Henrickson, H.H. Weetall, and B. Robertson. 2001. Одновременное обнаружение нескольких аналитов с порами нанометрового размера. Анальный. хим. 73:2268–2272. [PubMed] [Академия Google]
Weetall, and B. Robertson. 2001. Одновременное обнаружение нескольких аналитов с порами нанометрового размера. Анальный. хим. 73:2268–2272. [PubMed] [Академия Google]
10. Kasianowicz, J.J., S.E. Henrickson, M. Misakian, H.H. Weetall, B. Robertson, and V. Stanford. 2002. Физика ДНК, проходящей через нанометровые поры, и приложения для одновременного обнаружения нескольких аналитов. В Структура и динамика замкнутых полимеров. Дж. Дж. Касьянович, М. Келлермайер и Д. В. Димер, редакторы. Kluwer Academic Publishers, Дордрехт, Нидерланды. 141–163.
11. Бэйли Х. и Л. Джаясингхе. 2004. Функционально-инженерные каналы и поры (обзор). Мол. член биол. 21:209–220. [PubMed] [Google Scholar]
12. Li, J., D. Stein, C. McMullan, D. Branton, M.J. Aziz, and J.A. Golovchenko. 2001. Ионно-лучевое моделирование в нанометровых масштабах. Природа. 412: 166–169. [PubMed] [Google Scholar]
13. Harrell, C.C., S.B. Lee, and C.R. Martin. 2003. Синтетические мембраны с одинарными нанопорами и нанотрубками. Анальный. хим. 75:6861–6867. [PubMed] [Google Scholar]
Анальный. хим. 75:6861–6867. [PubMed] [Google Scholar]
14. Storm, A.J., JH Chen, X.S. Ling, H.W. Zandbergen, and C. Dekker. 2003. Изготовление твердотельных нанопор с точностью до одного нанометра. Нац. Матер. 2: 537–540. [PubMed] [Академия Google]
15. Yang, J., F. Lu, L.W. Kostiuk, and D.Y. Kwok. 2003. Электрокинетическая микроканальная батарея с помощью электрокинетических и микрожидкостных явлений. Дж. Микромех. Микроангл. 13:963–970. [Google Scholar]
16. Adcock, C., G.R. Smith, and M.S.P. Sansom. 1998. Электростатика и ионная селективность лиганд-управляемых каналов. Биофиз. Дж. 75:1211–1222. [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Wilson, GG, JM Pascual, N. Brooijmans, D. Murray, and A. Karlin. 2000. Собственный электростатический потенциал и промежуточное кольцо заряда в канале ацетилхолинового рецептора. J. Gen. Physiol. 115:93–106. [Бесплатная статья PMC] [PubMed] [Google Scholar]
18. Керамидас, А., А. Дж. Мурхаус, П. Р. Питер и П. Х. Барри. 2004. Лиганд-управляемые ионные каналы: механизмы, лежащие в основе ионной селективности. прог. Биофиз. Мол. биол. 86:161–204. [PubMed] [Google Scholar]
Питер и П. Х. Барри. 2004. Лиганд-управляемые ионные каналы: механизмы, лежащие в основе ионной селективности. прог. Биофиз. Мол. биол. 86:161–204. [PubMed] [Google Scholar]
19. Носков С. Ю., В. Им и Б. Ру. 2004. Проникновение ионов через канал α -гемолизин: теоретические исследования, основанные на броуновской динамике и теории электродиффузии Пуассона-Нернста-Планка. Биофиз. Дж. 87:2299–2309. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Jordan, PC 2005. Пятьдесят лет прогресса в исследованиях ионных каналов. IEEE транс. Нанобиология. 4:3–9. [PubMed] [Google Scholar]
21. Бхакди С. и Дж. Транум-Дженсен. 1991. Альфа-токсин Staphylococcus Aureus . микробиол. Откр. 55:733–751. [Бесплатная статья PMC] [PubMed] [Google Scholar]
22. Gouaux, JE, O. Braha, M.R. Hobaugh, L.Z. Song, S. Cheley, C. Shustak, and H. Bayley. 1994. Стехиометрия субъединиц Staphylococcal 9.0005 α -гемолизин в кристаллах и на мембранах: гептамерная трансмембранная пора. проц. Натл. акад. науч. США. 91:12828–12831. [Бесплатная статья PMC] [PubMed] [Google Scholar]
проц. Натл. акад. науч. США. 91:12828–12831. [Бесплатная статья PMC] [PubMed] [Google Scholar]
23. Сонг, Л. З., М. Р. Хобо, К. Шустак, С. Чели, Х. Бейли и Дж. Э. Гуо. 1996. Структура стафилококкового α -гемолизина, гептамерная трансмембранная пора. Наука. 274: 1859–1866. [PubMed] [Google Scholar]
24. Красильников О.В., Мерзляк П.Г., Юлдашева Л.Н., Родригес К.Г., Бхакди С., Валева А. 2000. Электрофизиологические доказательства гептамерной стехиометрии ионных каналов, образованных Staphylococcus aureus α -токсин в плоских липидных бислоях. Мол. микробиол. 37:1372–1378. [PubMed] [Google Scholar]
25. Красильников О.В., Сабиров Р.З., Терновский В.И., Мерзляк П.Г., Ташмухамедов Б.А. 1988. Структура Staphylococcus aureus α -индуцированного токсином ионного канала. Ген. физиол. Биофиз. 7: 467–473. [PubMed] [Google Scholar]
26. Красильников О.В. 2002. Проклейка каналов нейтральными полимерами. В Структура и динамика замкнутых полимеров. Дж. Дж. Касьянович, М. С. З. Келлермайер и Д. В. Димер, редакторы. Kluwer Academic Publishers, Дордрехт, Нидерланды. 97–115.
Дж. Дж. Касьянович, М. С. З. Келлермайер и Д. В. Димер, редакторы. Kluwer Academic Publishers, Дордрехт, Нидерланды. 97–115.
27. Füssle, R., S. Bhakdi, A. Sziegoleit, J. Tranum-Jensen, T. Kranz и H.J. Wellensiek. 1981. О механизме повреждения мембран Staphylococcus aureus α -токсином. Дж. Клеточная биология. 91:83–94. [бесплатная статья PMC] [PubMed] [Google Scholar]
28. Красильников О.В., Сабиров Р.З. 1989. Транспорт ионов через каналы, образованные в липидных бислоях Staphylococcus aureus α -токсином. Ген. физиол. Биофиз. 8: 213–222. [PubMed] [Академия Google]
29. Howorka, S., L. Movileanu, X.F. Lu, M. Magnon, S. Cheley, O. Braha и H. Bayley. 2000. Белковая пора с единственной полимерной цепью, связанной внутри просвета. Варенье. хим. соц. 122:2411–2416. [Google Scholar]
30. Мерзляк П.Г., Юлдашева Л.Н., Родригес К.Г., Карнейро К.М.М., Красильников О.В., Безруков С.М. 1999. Полимерные неэлектролиты для исследования геометрии пор: Применение к трансмембранному каналу α -токсина. Биофиз. Дж. 77:3023–3033. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Биофиз. Дж. 77:3023–3033. [Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Мовиляну Л., С. Чели, С. Ховорка, О. Браха и Х. Бейли. 2001. Локализация сужения в просвете трансмембранной поры путем направленного ковалентного присоединения полимерных молекул. J. Gen. Physiol. 117: 239–251. [PMC free article] [PubMed] [Google Scholar]
32. Красильников О.В., Терновский В.И., Ташмухамедов Б.А. 1981. Свойства α -стафилотоксин-индуцированных каналов проводимости в двухслойных фосфолипидных мембранах. Биофизика. 26:271–276. [PubMed] [Академия Google]
33. Менестрина Г. 1986. Ионные каналы, образованные токсином Staphylococcus aureus α : потенциалзависимое ингибирование двухвалентными и трехвалентными катионами. Дж. Член. биол. 90:177–190. [PubMed] [Google Scholar]
34. Красильников О.В., Терновский В.И., Сабиров Р.З., Зарипова Р.К., Ташмухамедов Б.А. 1986. Катион-анионная селективность каналов стафилотоксина в липидном бислое. Биофизика. 31: 606–610. [PubMed] [Google Scholar]
31: 606–610. [PubMed] [Google Scholar]
35. Мисакян М. и Дж. Дж. Касьянович. 2003. Электростатическое влияние на транспорт ионов через α канал HL. Дж. Член. биол. 195:137–146. [PubMed] [Google Scholar]
36. Носков С.Ю., Бернеш С., Ру Б. 2004. Контроль ионной селективности в калиевых каналах электростатическими и динамическими свойствами карбонильных лигандов. Природа. 431:830–834. [PubMed] [Google Scholar]
37. Безруков С.М., Водяной И., Брутян Р.А., Касьянович Дж.Дж. 1996. Динамика и свободная энергия разделения полимеров на наноразмерные поры. Макромолекулы. 29:8517–8522. [Академия Google]
38. Безруков С.М., Касьянович Дж.Дж. 2002. Динамическое разделение нейтральных полимеров на единый ионный канал. В Структура и динамика замкнутых полимеров. Дж. Дж. Касьянович, М. С. З. Келлермайер и Д. В. Димер, редакторы. Kluwer Academic Publishers, Дордрехт, Нидерланды. 117–130.
39. Красильников О.В., Безруков С.М. 2004. Разделение полимеров из неидеальных растворов на белковые пустоты. Макромолекулы. 37:2650–2657. [Академия Google]
Макромолекулы. 37:2650–2657. [Академия Google]
40. Касьянович, Дж. Дж. 2004. Нанопоры: зубная нить с ДНК. Нац. Матер. 3: 355–356. [PubMed] [Google Scholar]
41. Безруков С.М., Красильников О.В., Юлдашева Л.Н., Бережковский А.М., Родригес К.Г. 2004. Полезависимое влияние краун-эфира (18-краун-6) на ионную проводимость каналов α -гемолизина. Биофиз. Дж. 87:3162–3171. [Бесплатная статья PMC] [PubMed] [Google Scholar]
42. Безруков С.М., Касьянович Дж.Дж. 1997. Состояние заряда ионного канала контролирует проникновение нейтрального полимера в его поры. Евро. Биофиз. Дж. 26:471–476. [PubMed] [Академия Google]
43. Акабас, М. Х., Д. А. Штауффер, М. Сюй и А. Карлин. 1992. Исследована структура канала ацетилхолинового рецептора у мутантов с заменой цистеина. Наука. 258:307–310. [PubMed] [Google Scholar]
44. Монталь М. и П. Мюллер. 1972. Формирование бимолекулярных мембран из липидных монослоев и изучение их электрических свойств. проц. Натл. акад. науч. США. 69:3561–3566. [PMC free article] [PubMed] [Google Scholar]
акад. науч. США. 69:3561–3566. [PMC free article] [PubMed] [Google Scholar]
45. Красильников О.В., Мерзляк П.Г., Юлдашева Л.Н., Ногейра Р.А., Родригес К.Г. 1995. Нестохастическое распределение одиночных каналов в плоских липидных бислоях. Биохим. Биофиз. Акта. 1233: 105–110. [PubMed] [Google Scholar]
46. Барри П. Х. и Дж. В. Линч. 1991. Потенциалы жидкостного соединения и эффекты мелких клеток в анализе пэтч-клэмп. Дж. Член. биол. 121:101–117. [PubMed] [Google Scholar]
47. Нг, Б. и П. Х. Барри. 1995. Измерение ионной проводимости и подвижности некоторых менее распространенных органических ионов, необходимых для коррекции потенциала перехода в электрофизиологии. Дж. Нейроски. Методы. 56:37–41. [PubMed] [Академия Google]
48. Валева А., Дж. Понгс, С. Бхакди и М. Палмер. 1997. Стафилококковый α -токсин: роль N-конца в формировании гептамерной поры — флуоресцентное исследование. Биохим. Биофиз. Акта. 1325: 281–286. [PubMed] [Google Scholar]
49. Guex, N., and M.C. Peitsch. 1997. SWISS-MODEL и Swiss-PdbViewer: среда для сравнительного моделирования белков. Электрофорез. 18:2714–2723. [PubMed] [Google Scholar]
Guex, N., and M.C. Peitsch. 1997. SWISS-MODEL и Swiss-PdbViewer: среда для сравнительного моделирования белков. Электрофорез. 18:2714–2723. [PubMed] [Google Scholar]
50. Neumcke, B. 1970. Поток ионов через липидные бислойные мембраны с заряженными поверхностями. Биофизика. 6: 231–240. [PubMed] [Академия Google]
51. Маркин В.С., Чисмажев Ю.А. 1974. Индуцированный ионный транспорт. Наука, Москва.
52. Белл, Дж. Э. и К. Миллер. 1984. Влияние поверхностного заряда фосфолипидов на ионную проводимость в канале K + саркоплазматического ретикулума. Биофиз. Дж. 45:279–287. [PMC free article] [PubMed] [Google Scholar]
53. Красильников О.В., Капистрано М.Ф.П., Юлдашева Л.Н., Ногейра Р.А. 1997. Влияние токсина Cys-130 S-aureus α на плоский липидный бислой и мембраны эритроцитов. Дж. Член. биол. 156: 157–172. [PubMed] [Академия Google]
54. Красильников О.В., Мерзляк П.Г., Сабиров Р.З., Терновский В.И., Зарипова Р.К. 1988. Влияние pH на потенциал-зависимость каналов стафилококкового токсина, функционирующих в фосфатидилхолиновом бислое. Укр. Биохим. ж. 60:60–66. [PubMed] [Google Scholar]
Укр. Биохим. ж. 60:60–66. [PubMed] [Google Scholar]
55. Красильников О.В., Юлдашева Л.Н., Мерзляк П.Г., Капистрано М.Ф.П., Ногейра Р.А. 1997. Шарнирная часть -токсина S. aureus α пересекает липидный бислой и является частью транс -устье канала. Биохим. Биофиз. Акта. 1329: 51–60. [PubMed] [Google Scholar]
56. Корчев Ю.Е., Башфорд К.Л., Алдер Г.М., Касьянович Дж., Пастернак К.А. 1995. Низкопроводящие состояния одноионного канала не закрыты. Дж. Член. биол. 147: 233–239. [PubMed] [Google Scholar]
57. Красильников О.В., Мерзляк П.Г., Юлдашева Л.Н., Родригес К.Г., Ногейра Р.А. 1999. Влияние гепарина на α -стафилотоксин образованный канал. Биохим. Биофиз. Акта. 1417: 167–182. [PubMed] [Академия Google]
58. Красильников О.В., Мерзляк П.Г., Юлдашева Л.Н., Ногейра Р.А. 1998. Эксперименты по определению размера канала в многоканальных бислоях. Ген. физиол. Биофиз. 17:349–363. [PubMed] [Google Scholar]
59. Krekel, F., A. K. Samland, P. Maceroux, N. Amrhein, and JNS Evans. 2000. Определение значения pK(a) C115 в MurA (UDP- N -ацетилглюкозаминэнолпирувилтрансфераза) из Enterobacter cloacae . Биохимия. 39:12671–12677. [PubMed] [Академия Google]
K. Samland, P. Maceroux, N. Amrhein, and JNS Evans. 2000. Определение значения pK(a) C115 в MurA (UDP- N -ацетилглюкозаминэнолпирувилтрансфераза) из Enterobacter cloacae . Биохимия. 39:12671–12677. [PubMed] [Академия Google]
60. Валева, А., А. Вайссер, Б. Уокер, М. Кехо, Х. Бэйли, С. Бхакди и М. Палмер. 1996. Молекулярная архитектура токсиновой поры: последовательность из 15 остатков выстилает трансмембранный канал стафилококкового α -токсина. EMBO J. 15: 1857–1864. [PMC free article] [PubMed] [Google Scholar]
АТФ-высвобождающий Maxi-Cl канал: его сущность, молекулярные партнеры и физиологические/патофизиологические последствия
1. Замараева М.В., Сабиров Р.З., Маэно Э., Андо- Акацука Ю., Бессонова С.В., Окада Ю. Клетки погибают с повышенным содержанием цитозольного АТФ во время апоптоза: исследование биолюминесценции с внутриклеточной люциферазой. Смерть клеток 2005;12:1390–1397. doi: 10.1038/sj.cdd.4401661. [PubMed] [CrossRef] [Google Scholar]
2. Kennedy H.J., Pouli A.E., Ainscow E.K., Jouaville L.S., Rizzuto R., Rutter G.A. Глюкоза генерирует микродомены АТФ субплазматической мембраны в одиночных островковых бета-клетках. Потенциальная роль стратегически расположенных митохондрий. Дж. Биол. хим. 1999; 274:13281–13291. doi: 10.1074/jbc.274.19.13281. [PubMed] [CrossRef] [Google Scholar]
Kennedy H.J., Pouli A.E., Ainscow E.K., Jouaville L.S., Rizzuto R., Rutter G.A. Глюкоза генерирует микродомены АТФ субплазматической мембраны в одиночных островковых бета-клетках. Потенциальная роль стратегически расположенных митохондрий. Дж. Биол. хим. 1999; 274:13281–13291. doi: 10.1074/jbc.274.19.13281. [PubMed] [CrossRef] [Google Scholar]
3. Gribble F.M., Loussouarn G., Tucker S.J., Zhao C., Nichols C.G., Ashcroft F.M. Новый метод измерения субмембранной концентрации АТФ. Дж. Биол. хим. 2000;275:30046–30049. doi: 10.1074/jbc.M001010200. [PubMed] [CrossRef] [Google Scholar]
4. Замараева М.В., Сабиров Р.З., Манабе К., Окада Ю. Активация Ca(2+)-зависимого гликолиза опосредует апоптотическое повышение АТФ в клетках HeLa. Биохим. Биофиз. Рез. коммун. 2007; 363: 687–693. doi: 10.1016/j.bbrc.2007.09.019. [PubMed] [CrossRef] [Google Scholar]
5. Брейк А.Дж., Джулиус Д. Передача сигналов внеклеточными нуклеотидами. Анну. Преподобный Cell Dev. биол. 1996; 12: 519–541. doi: 10.1146/annurev.cellbio.12.1.519. [PubMed] [CrossRef] [Академия Google]
doi: 10.1146/annurev.cellbio.12.1.519. [PubMed] [CrossRef] [Академия Google]
6. Окада Ю., Окада Т., Ислам М.Р., Сабиров Р.З. Молекулярная идентичность и активность высвобождения АТФ двух типов анионных каналов, регулирующих объем, VSOR и Maxi-Cl. Курс. Верхний. член 2018; 81: 125–176. doi: 10.1016/bs.ctm.2018.07.004. [PubMed] [CrossRef] [Google Scholar]
7. Циммерманн Х. Два новых семейства эктонуклеотидаз: молекулярные структуры, каталитические свойства и поиск функции. Тренды Фармакол. науч. 1999; 20: 231–236. doi: 10.1016/S0165-6147(99)01293-6. [PubMed] [CrossRef] [Академия Google]
8. Линден Дж., Кох-Нольте Ф., Даль Г. Высвобождение пуринов, метаболизм и передача сигналов при воспалительной реакции. Анну. Преподобный Иммунол. 2019;37:325–347. doi: 10.1146/annurev-иммунол-051116-052406. [PubMed] [CrossRef] [Google Scholar]
9. Дуайер К.М., Кишор Б.К., Робсон С.К. Преобразование внеклеточного АТФ в аденозин: главный переключатель в почечной патологии и здоровье. Нац. Преподобный Нефрол. 2020;16:509–524. doi: 10.1038/s41581-020-0304-7. [PubMed] [CrossRef] [Google Scholar]
Нац. Преподобный Нефрол. 2020;16:509–524. doi: 10.1038/s41581-020-0304-7. [PubMed] [CrossRef] [Google Scholar]
10. Бернсток Г. Физиология и патофизиология пуринергической нейротрансмиссии. Физиол. Ред. 2007; 87: 659–797. doi: 10.1152/physrev.00043.2006. [PubMed] [CrossRef] [Google Scholar]
11. Бернсток Г. Пуринергическая передача сигналов: непопулярное начало, признание и захватывающее будущее. Биоэссе. 2012; 34: 218–225. doi: 10.1002/bies.201100130. [PubMed] [CrossRef] [Google Scholar]
12. Сабиров Р.З., Окада Ю. АТФ-проводящий макси-анионный канал: новый игрок в стресс-сенсорной трансдукции. Япония. Дж. Физиол. 2004; 54:7–14. doi: 10.2170/jjphysiol.54.7. [PubMed] [CrossRef] [Академия Google]
13. Сабиров Р.З., Окада Ю. Высвобождение АТФ по анионным каналам. Пуринергический сигнал. 2005; 1: 311–328. doi: 10.1007/s11302-005-1557-0. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
14. Сабиров Р.З., Окада Ю. Макси-анионный канал: классический канал, играющий новые роли через неопознанную молекулярную сущность. Дж. Физиол. науч. 2009; 59:3–21. doi: 10.1007/s12576-008-0008-4. [PubMed] [CrossRef] [Google Scholar]
Дж. Физиол. науч. 2009; 59:3–21. doi: 10.1007/s12576-008-0008-4. [PubMed] [CrossRef] [Google Scholar]
15. Li A., Banerjee J., Leung C.T., Peterson-Yantorno K., Stamer W.D., Civan M.M. Механизмы высвобождения АТФ, активизирующий этап пуринергической динамики. Клетка. Физиол. Биохим. 2011; 28:1135–1144. дои: 10.1159/000335865. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
16. Лазаровский Э.Р., Буше Р.К., Харден Т.К. Механизмы высвобождения нуклеотидов и интеграция их действия в качестве молекул, активирующих P2X- и P2Y-рецепторы. Мол. Фармакол. 2003; 64: 785–795. doi: 10.1124/мол.64.4.785. [PubMed] [CrossRef] [Google Scholar]
17. Лазаровский Э.Р., Сесма Дж.И., Семинарио-Видаль Л., Креда С.М. Молекулярные механизмы высвобождения пуриновых и пиримидиновых нуклеотидов. Доп. Фармакол. 2011;61:221–261. дои: 10.1016/B978-0-12-385526-8.00008-4. [PubMed] [CrossRef] [Google Scholar]
18. Лазаровский Э.Р. Везикулярный и кондуктивный механизмы высвобождения нуклеотидов. Пуринергический сигнал. 2012; 8: 359–373. doi: 10.1007/s11302-012-9304-9. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Пуринергический сигнал. 2012; 8: 359–373. doi: 10.1007/s11302-012-9304-9. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
19. Таруно А., Вингтдо В., Омото М., Ма З., Дворянчиков Г., Ли А., Адриен Л., Чжао Х. , Леунг С., Абернети М. и др. Ионный канал CALHM1 опосредует пуринергическую нейротрансмиссию сладкого, горького вкуса и вкуса умами. Природа. 2013;495: 223–226. doi: 10.1038/nature11906. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
20. Сандерсон Дж., Дартт Д.А., Тринкаус-Рэндалл В., Пинтор Дж., Сиван М.М., Деламер Н.А., Флетчер Э.Л., Солт Т.Е., Гроше А. ., Митчелл Ч.Х. Пурины в глазах: недавние доказательства физиологической и патологической роли пуринов в RPE, нейронах сетчатки, астроцитах, клетках Мюллера, хрусталике, трабекулярной сети, роговице и слезной железе. Эксп. Глаз Res. 2014; 127: 270–279. doi: 10.1016/j.exer.2014.08.009. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
21. Морияма Ю., Хиаса М., Сакамото С. , Омотэ Х., Номура М. Транспортер везикулярных нуклеотидов (ВНУТ): Внешний вид актрисы на стадия пуринергической сигнализации. Пуринергический сигнал. 2017;13:387–404. doi: 10.1007/s11302-017-9568-1. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
, Омотэ Х., Номура М. Транспортер везикулярных нуклеотидов (ВНУТ): Внешний вид актрисы на стадия пуринергической сигнализации. Пуринергический сигнал. 2017;13:387–404. doi: 10.1007/s11302-017-9568-1. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
22. Таруно А. Каналы выпуска ATP. Междунар. Дж. Мол. науч. 2018;19:808. doi: 10.3390/ijms1
08. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
23. Praetorius H.A., Leipziger J. Высвобождение АТФ из невозбудимых клеток. Пуринергический сигнал. 2009; 5: 433–446. doi: 10.1007/s11302-009-9146-2. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
24. Vallon V., Unwin R., Inscho E.W., Leipziger J., Kishore B.K. Внеклеточные нуклеотиды и рецепторы P2 в функции почек. Физиол. 2020; 100:211–269. doi: 10.1152/physrev.00038.2018. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
25. Lohman A.W., Isakson B.E. Дифференциация коннексиновых полуканалов и паннексиновых каналов при высвобождении клеточного АТФ. ФЭБС лат. 2014;588:1379–1388. doi: 10.1016/j.febslet.2014.02.004. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
ФЭБС лат. 2014;588:1379–1388. doi: 10.1016/j.febslet.2014.02.004. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
26. Юнг А.К., Патил К.С., Джексон М.Ф. Паннексин-1 в ЦНС: новые концепции здоровья и болезней. Дж. Нейрохим. 2020; 154: 468–485. doi: 10.1111/jnc.15004. [PubMed] [CrossRef] [Google Scholar]
27. Acosta M.L., Mat Nor M.N., Guo C.X., Mugisho O.O., Coutinho F.P., Rupenthal I.D., Green CR. Терапия коннексином: блокирование пор гемиканала коннексином отличается от блокирования каналов паннексина или щели. соединения. Нейронная регенерация. Рез. 2021; 16: 482–488. дои: 10.4103/1673-5374.2. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
28. Navis K.E., Fan C.Y., Trang T., Thompson R.J., Derksen D.J. Каналы паннексина 1 как терапевтическая мишень: структура, торможение и перспективы. АКС хим. Неврологи. 2020;11:2163–2172. doi: 10.1021/acschemneuro.0c00333. [PubMed] [CrossRef] [Google Scholar]
29. Андреев Р., Оливейра-Джакомелли А. , Рибейро Д.Э., Глейзер Т., Арно-Сампайо В.Ф., Ламеу К., Ульрих Х. Рецептор P2X7: центральный узел заболевания головного мозга. Фронт. Мол. Неврологи. 2020;13:124. дои: 10.3389/fnmol.2020.00124. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
, Рибейро Д.Э., Глейзер Т., Арно-Сампайо В.Ф., Ламеу К., Ульрих Х. Рецептор P2X7: центральный узел заболевания головного мозга. Фронт. Мол. Неврологи. 2020;13:124. дои: 10.3389/fnmol.2020.00124. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
30. Panicucci C., Raffaghello L., Bruzzone S., Baratto S., Principi E., Minetti C., Gazzerro E., Bruno C. Ось eATP / P2X7R: организованный путь, запускающий активацию воспаления при мышечных заболеваниях. Междунар. Дж. Мол. науч. 2020;21:5963. doi: 10.3390/ijms21175963. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
31. Ma Z., Tanis J.E., Taruno A., Foskett J.K. Ионные каналы модулятора гомеостаза кальция (CALHM). Пфлуг. Арка 2016;468:395–403. doi: 10.1007/s00424-015-1757-6. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
32. Wei L., Mousawi F., Li D., Roger S., Li J., Yang X., Jiang L.H. Высвобождение аденозинтрифосфата и P2 передача сигналов рецептора в зависимой от канала Piezo1 механорегуляции.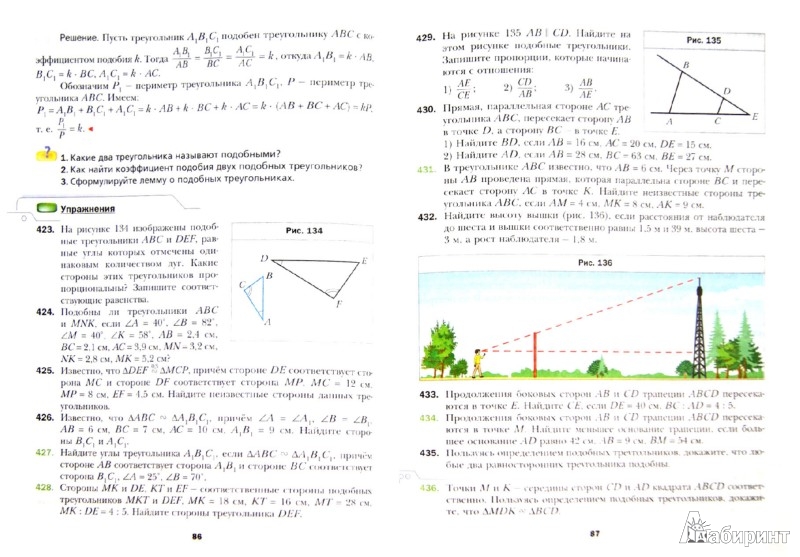 Фронт. Фармакол. 2019;10:1304. doi: 10.3389/fphar.2019.01304. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Фронт. Фармакол. 2019;10:1304. doi: 10.3389/fphar.2019.01304. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
33. Сабиров Р.З., Мерзляк П.Г., Ислам М.Р., Окада Т., Окада Ю. Свойства, функции и патофизиология максианионных каналов. Пфлуг. Арка 2016; 468:405–420. doi: 10.1007/s00424-015-1774-5. [PubMed] [CrossRef] [Академия Google]
34. Элорза-Видаль X., Гайтан-Пенас Х., Эстевес Р. Хлоридные каналы в астроцитах: структура, роль в гомеостазе головного мозга и последствия при заболеваниях. Междунар. Дж. Мол. науч. 2019;20:1034. doi: 10.3390/ijms20051034. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
35. Окада Ю., Окада Т., Сато-Нумата К., Ислам М.Р., Андо-Акацука Ю., Нумата Т., Кубо М., Симидзу Т., Курбанназарова Р.С., Марунака Ю. и др. Активируемые по объему клетки и коррелированные по объему анионные каналы в клетках млекопитающих: их биофизические, молекулярные и фармакологические свойства. Фармакол. Версия 2019 г.;71:49–88. doi: 10.1124/пр. 118.015917. [PubMed] [CrossRef] [Google Scholar]
118.015917. [PubMed] [CrossRef] [Google Scholar]
36. Окада Ю., Сабиров Р.З., Сато-Нумата К., Нумата Т. Индукция и защита клеток от гибели путем активации повсеместно экспрессируемых анион/катионных каналов. Часть 1: Роль VSOR/VRAC в регуляции объема клеток, высвобождении обоюдоострых сигналов и апоптотической/некротической гибели клеток. Фронт. Сотовый Дев. биол. 2021;8:614040. doi: 10.3389/fcell.2020.614040. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
37. Дженч Т.Дж., Пуш М. Хлоридные каналы и транспортеры CLC: структура, функция, физиология и заболевание. Физиол. Ред. 2018; 98: 1493–1590. doi: 10.1152/physrev.00047.2017. [PubMed] [CrossRef] [Google Scholar]
38. Блаунт П., Искла И. Жизнь с бактериальными механочувствительными каналами, от открытия до физиологии и фармакологической цели. микробиол. Мол. биол. 2020;84:e00055-00019. doi: 10.1128/MMBR.00055-19. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
39. Blatz A.L., Magleby K. L. Одиночные потенциалзависимые хлорид-селективные каналы большой проводимости в культивируемых мышцах крыс. Биофиз. Дж. 1983;43:237–241. doi: 10.1016/S0006-3495(83)84344-6. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
L. Одиночные потенциалзависимые хлорид-селективные каналы большой проводимости в культивируемых мышцах крыс. Биофиз. Дж. 1983;43:237–241. doi: 10.1016/S0006-3495(83)84344-6. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
40. Riquelme G., Stutzin A., Barros L.F., Liberona J.L. Хлоридный канал из плаценты человека, преобразованный в гигантские липосомы. Являюсь. Дж. Обст. Гинекол. 1995; 173: 733–738. doi: 10.1016/0002-9378(95)
-1. [PubMed] [CrossRef] [Google Scholar]
41. Стрэндж К., Эмма Ф., Джексон П.С. Клеточная и молекулярная физиология объемочувствительных анионных каналов. Являюсь. Дж. Физиол. 1996;270:С711–С730. doi: 10.1152/ajpcell.1996.270.3.C711. [PubMed] [CrossRef] [Google Scholar]
42. Riquelme G. Апикальный максихлоридный канал из плаценты человека: 12 лет после первых электрофизиологических записей. биол. Рез. 2006; 39: 437–445. doi: 10.4067/S0716-97602006000300006. [PubMed] [CrossRef] [Google Scholar]
43. Bell PD, Komlosi P. , Zhang Z.R. АТФ как медиатор передачи сигналов клетками плотного пятна. Пуринергический сигнал. 2009; 5: 461–471. doi: 10.1007/s11302-009-9148-0. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
, Zhang Z.R. АТФ как медиатор передачи сигналов клетками плотного пятна. Пуринергический сигнал. 2009; 5: 461–471. doi: 10.1007/s11302-009-9148-0. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
44. Riquelme G. Плацентарные хлоридные каналы: обзор. Плацента. 2009; 30: 659–669. doi: 10.1016/j.placenta.2009.06.002. [PubMed] [CrossRef] [Google Scholar]
45. O’Donnell M.J., Kelly S.P., Nurse C.A., Wood C.M. Макси-канал Cl(-) в культивируемых клетках мостовой из жабр пресноводной радужной форели Oncorhynchus mykiss. Дж. Эксп. биол. 2001; 204: 1783–1794. дои: 10.1242/jeb.204.10.1783. [PubMed] [CrossRef] [Google Scholar]
46. Woll K.H., Leibowitz MD, Neumcke B., Hille B. Анионный канал с высокой проводимостью в скелетных мышцах взрослых амфибий. Пфлуг. Арка 1987;410:632–640. doi: 10.1007/BF00581324. [PubMed] [CrossRef] [Google Scholar]
47. Hussy N. Кальций-активированные хлоридные каналы в культивируемых эмбриональных спинномозговых нейронах Xenopus. Дж. Нейрофизиол. 1992;68:2042–2050. doi: 10.1152/jn.1992.68.6.2042. [PubMed] [CrossRef] [Google Scholar]
Дж. Нейрофизиол. 1992;68:2042–2050. doi: 10.1152/jn.1992.68.6.2042. [PubMed] [CrossRef] [Google Scholar]
48. Stumpff F., Martens H., Bilk S., Aschenbach J.R., Gabel G. Культивируемые эпителиальные клетки рубца экспрессируют канал с большой проводимостью, проницаемый для хлорида, бикарбоната и ацетата. . Пфлуг. Арка 2009 г.;457:1003–1022. doi: 10.1007/s00424-008-0566-6. [PubMed] [CrossRef] [Google Scholar]
49. Stumpff F., Georgi M.I., Mundhenk L., Rabbani I., Fromm M., Martens H., Gunzel D. Первичные культуры рубца овец и omasum и исходный эпителий: Барьерная функция связана с экспрессией белков плотных контактов. Дж. Эксп. биол. 2011; 214:2871–2882. doi: 10.1242/jeb.055582. [PubMed] [CrossRef] [Google Scholar]
50. Георги М.И., Розендаль Дж., Эрнст Ф., Гюнцель Д., Ашенбах Дж.Р., Мартенс Х., Штумпф Ф. анионные каналы, проницаемые для анионов короткоцепочечных жирных кислот. Пфлуг. Арка 2014;466:1689–1712. doi: 10.1007/s00424-013-1386-x. [PubMed] [CrossRef] [Google Scholar]
51.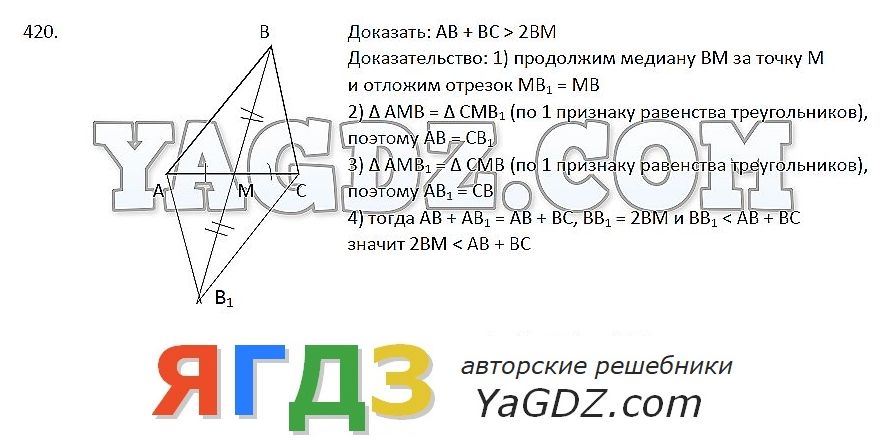 Stumpff F. Взгляд на вонючую сторону физиологии: транспорт короткоцепочечных жирных кислот. Пфлуг. Арка 2018; 470: 571–598. doi: 10.1007/s00424-017-2105-9. [PubMed] [CrossRef] [Google Scholar]
Stumpff F. Взгляд на вонючую сторону физиологии: транспорт короткоцепочечных жирных кислот. Пфлуг. Арка 2018; 470: 571–598. doi: 10.1007/s00424-017-2105-9. [PubMed] [CrossRef] [Google Scholar]
52. Groschner K., Kukovetz W.R. Потенциально-чувствительные хлоридные каналы большой проводимости в мембране эндотелиальных клеток аорты свиньи. Пфлуг. Арка 1992; 421: 209–217. doi: 10.1007/BF00374829. [PubMed] [CrossRef] [Академия Google]
53. Олесен С.П., Бундгаард М. Хлорид-селективные каналы большой проводимости в эндотелиальных клетках бычьей аорты. Акта Физиол. Сканд. 1992; 144: 191–198. doi: 10.1111/j.1748-1716.1992.tb09285.x. [PubMed] [CrossRef] [Google Scholar]
54. Сабиров Р.З., Дутта А.К., Окада Ю. Объемно-зависимый АТФ-проводящий анионный канал большой проводимости как путь индуцированного набуханием высвобождения АТФ. J. Gen. Physiol. 2001; 118: 251–266. doi: 10.1085/jgp.118.3.251. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
55. Ислам М. Р., Урамото Х., Окада Т., Сабиров Р.З., Окада Ю. Макси-анионный канал и полуканал паннексина 1 представляют собой отдельные пути индуцированного набуханием высвобождения АТФ в мышиных клетках фибросаркомы L929. Являюсь. Дж. Физиол. Клеточная физиол. 2012; 303:C924–C935. doi: 10.1152/ajpcell.00459.2011. [PubMed] [CrossRef] [Google Scholar]
Р., Урамото Х., Окада Т., Сабиров Р.З., Окада Ю. Макси-анионный канал и полуканал паннексина 1 представляют собой отдельные пути индуцированного набуханием высвобождения АТФ в мышиных клетках фибросаркомы L929. Являюсь. Дж. Физиол. Клеточная физиол. 2012; 303:C924–C935. doi: 10.1152/ajpcell.00459.2011. [PubMed] [CrossRef] [Google Scholar]
56. Bell P.D., Lapointe J.Y., Sabirov R., Hayashi S., Peti-Peterdi J., Manabe K., Kovacs G., Okada Y. Передача сигналов клетками плотной макулы включает Высвобождение АТФ через макси-анионный канал. проц. Натл. акад. науч. США. 2003; 100:4322–4327. doi: 10.1073/pnas.0736323100. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
57. Дутта А.К., Сабиров Р.З., Урамото Х., Окада Ю. Роль АТФ-проводящего анионного канала в высвобождении АТФ из кардиомиоцитов новорожденных крыс в условиях ишемии или гипоксии. Дж. Физиол. 2004; 559: 799–812. doi: 10.1113/jphysiol.2004.069245. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
58. Дутта А.К., Корчев Ю.Е., Шевчук А.И., Хаяши С., Окада Ю., Сабиров Р.З. Пространственное распределение макси-анионного канала на кардиомиоцитах, обнаруженное методом смарт-пластырей. Биофиз. Дж. 2008;94: 1646–1655. doi: 10.1529/biophysj.107.117820. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Дутта А.К., Корчев Ю.Е., Шевчук А.И., Хаяши С., Окада Ю., Сабиров Р.З. Пространственное распределение макси-анионного канала на кардиомиоцитах, обнаруженное методом смарт-пластырей. Биофиз. Дж. 2008;94: 1646–1655. doi: 10.1529/biophysj.107.117820. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
59. Лю Х.Т., Ташмухамедов Б.А., Иноуэ Х., Окада Ю., Сабиров Р.З. Роль двух типов анионных каналов в высвобождении глутамата из астроцитов мыши при ишемическом или осмотическом стрессе. Глия. 2006; 54: 343–357. doi: 10.1002/glia.20400. [PubMed] [CrossRef] [Google Scholar]
60. Liu H.T., Sabirov R.Z., Okada Y. Депривация кислорода и глюкозы индуцирует высвобождение АТФ через макси-анионные каналы в астроцитах. Пуринергический сигнал. 2008; 4: 147–154. doi: 10.1007/s11302-007-9077-8. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
61. Сабиров Р.З., Шейко Т., Лю Х., Дэн Д., Окада Ю., Крейген В.Дж. Генетическая демонстрация того, что максианионный канал и напряжение плазматической мембраны -зависимые анионные каналы являются неродственными белками. Дж. Биол. хим. 2006; 281:1897–1904. doi: 10.1074/jbc.M509482200. [PubMed] [CrossRef] [Google Scholar]
Дж. Биол. хим. 2006; 281:1897–1904. doi: 10.1074/jbc.M509482200. [PubMed] [CrossRef] [Google Scholar]
62. Тойчиев А.Х., Сабиров Р.З., Такахаши Н., Андо-Акацука Ю., Лю Х., Шинтани Т., Нода М., Окада Ю. Активация макси-аниона канала путем дефосфорилирования белка тирозина. Являюсь. Дж. Физиол. Клеточная физиол. 2009 г.;297:C990–C1000. doi: 10.1152/ajpcell.00131.2009. [PubMed] [CrossRef] [Google Scholar]
63. Курбанназарова Р.С., Бессонова С.В., Окада Ю., Сабиров Р.З. Анионные каналы, активируемые набуханием, необходимы для регуляции объема тимоцитов мыши. Междунар. Дж. Мол. науч. 2011;12:9125–9137. doi: 10.3390/ijms12129125. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
64. Schwarze W., Kolb H.A. Потенциал-зависимая кинетика анионного канала большой единицы проводимости в макрофагах и мембранах миотрубочек. Пфлуг. Арка 1984;402:281–291. doi: 10.1007/BF00585511. [PubMed] [CrossRef] [Google Scholar]
65. Гелетюк В.И., Казаченко В.Н. Одиночные каналы Cl (-) в нейронах моллюсков: множественность состояний проводимости. Дж. Член. биол. 1985; 86: 9–15. doi: 10.1007/BF01871605. [PubMed] [CrossRef] [Google Scholar]
Дж. Член. биол. 1985; 86: 9–15. doi: 10.1007/BF01871605. [PubMed] [CrossRef] [Google Scholar]
66. Sun X.P., Supplisson S., Mayer E. Хлоридные каналы в миоцитах толстой кишки кролика регулируются чувствительным к коклюшному токсину G-белком. Являюсь. Дж. Физиол. 1993; 264:G774–G785. doi: 10.1152/jpgi.1993.264.4.G774. [PubMed] [CrossRef] [Google Scholar]
67. Сабиров Р.З., Окада Ю. Широкие наноскопические поры максианионного канала соответствуют его функции АТФ-проводящего пути. Биофиз. Дж. 2004; 87: 1672–1685. doi: 10.1529/biophysj.104.043174. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
68. Hurnak O., Zachar J. Селективность максихлоридных каналов в линии мышечных клеток крыс L6. Ген. физиол. Биофиз. 1995; 14:91–105. [PubMed] [Google Scholar]
69. Soejima M., Kokubun S. Одиночный анионселективный канал и его ионная селективность в гладкомышечных клетках сосудов. Пфлуг. Арка 1988;411:304–311. doi: 10.1007/BF00585119. [PubMed] [CrossRef] [Google Scholar]
70.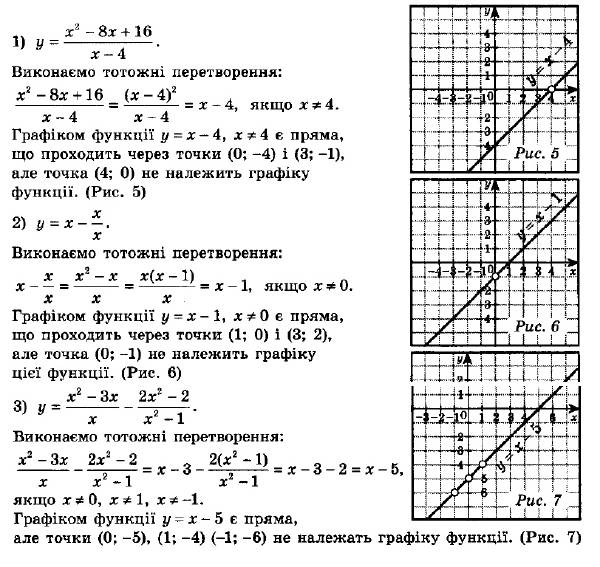 Дутта А.К., Окада Ю., Сабиров Р.З. Регуляция АТФ-проводящего анионного канала большой проводимости и высвобождение АТФ, вызванное набуханием, арахидоновой кислотой. Дж. Физиол. 2002; 542: 803–816. doi: 10.1113/jphysiol.2002.019802. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Дутта А.К., Окада Ю., Сабиров Р.З. Регуляция АТФ-проводящего анионного канала большой проводимости и высвобождение АТФ, вызванное набуханием, арахидоновой кислотой. Дж. Физиол. 2002; 542: 803–816. doi: 10.1113/jphysiol.2002.019802. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
71. Сабиров Р.З., Курбанназарова Р.С., Меланова Н.Р., Окада Ю. Объемочувствительные анионные каналы опосредуют осмочувствительное высвобождение глутатиона из тимоцитов крысы. ПЛОС ОДИН. 2013;8:e55646. doi: 10.1371/journal.pone.0055646. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
72. Сабиров Р.З., Мерзляк П.Г. Споры о Plasmalemmal VDAC и загадка макси-анионного канала. Биохим. Биофиз. Акта. 2012; 1818:1570–1580. doi: 10.1016/j.bbamem.2011.09.024. [PubMed] [CrossRef] [Google Scholar]
73. Falke LC, Misler S. Активность ионных каналов во время регуляции объема клональными клетками нейробластомы N1E115. проц. Натл. акад. науч. США. 1989; 86: 3919–3923. doi: 10.1073/pnas. 86.10.3919. [Статья PMC free] [PubMed] [CrossRef] [Google Scholar]
86.10.3919. [Статья PMC free] [PubMed] [CrossRef] [Google Scholar]
74. Jalonen T. Одноканальные характеристики анионного канала большой проводимости в астроцитах коры головного мозга крысы в первичной культуре. Глия. 1993;9:227–237. doi: 10.1002/glia.4400
. [PubMed] [CrossRef] [Google Scholar]
75. Schwiebert E.M., Mills J.W., Stanton B.A. Цитоскелет на основе актина регулирует хлоридный канал и объем клеток в клеточной линии собирательных трубочек почки. Дж. Биол. хим. 1994; 269:7081–7089. doi: 10.1016/S0021-9258(17)37249-6. [PubMed] [CrossRef] [Google Scholar]
76. Пети-Петерди Дж., Моришима С., Белл П.Д., Окада Ю. Флуоресцентная визуализация живого юкстагломерулярного аппарата с двухфотонным возбуждением. Являюсь. Дж. Физиол. Рен. Физиол. 2002;283:F197–F201. doi: 10.1152/ajprenal.00356.2001. [PubMed] [CrossRef] [Google Scholar]
77. Komlosi P., Fintha A., Bell P.D. Выяснение взаимосвязи между объемом клеток macula densa и концентрацией/осмоляльностью растворенного вещества в просвете. почки инт. 2006; 70: 865–871. doi: 10.1038/sj.ki.5001633. [PubMed] [CrossRef] [Google Scholar]
почки инт. 2006; 70: 865–871. doi: 10.1038/sj.ki.5001633. [PubMed] [CrossRef] [Google Scholar]
78. Kawahara K., Takuwa N. Бомбезин активирует хлоридные каналы большой проводимости в фибробластах Swiss 3T3. Биохим. Биофиз. Рез. коммун. 1991; 177: 292–298. дои: 10.1016/0006-291Х(91)
-Н. [PubMed] [CrossRef] [Google Scholar]
79. Bajnath R.B., Groot J.A., de Jonge HR, Kansen M., Bijman J. Ионофор кальция плюс иссечение индуцируют хлоридный канал большой проводимости в участках мембраны клеток карциномы толстой кишки человека HT -29кл.19А. Опыт. 1993; 49: 313–316. doi: 10.1007/BF01
9. [PubMed] [CrossRef] [Google Scholar]80. Dezaki K., Tsumura T., Maeno E., Okada Y. Опосредованное рецепторами облегчение регуляции клеточного объема за счет высвобождения АТФ, вызванного набуханием, в эпителиальных клетках человека. Япония. Дж. Физиол. 2000; 50: 235–241. doi: 10.2170/jjphysiol.50.235. [PubMed] [CrossRef] [Академия Google]
81. Kajita H., Kotera T., Shirakata Y. , Ueda S., Okuma M., Oda-Ohmae K., Takimoto M., Urade Y., Okada Y. Макси-канал Cl(-), связанный с Рецепторы эндотелина В в базолатеральной мембране париетальных клеток морской свинки. Дж. Физиол. 1995; 488: 65–75. doi: 10.1113/jphysiol.1995.sp020946. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
, Ueda S., Okuma M., Oda-Ohmae K., Takimoto M., Urade Y., Okada Y. Макси-канал Cl(-), связанный с Рецепторы эндотелина В в базолатеральной мембране париетальных клеток морской свинки. Дж. Физиол. 1995; 488: 65–75. doi: 10.1113/jphysiol.1995.sp020946. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
82. Schwiebert E.M., Karlson K.H., Friedman P.A., Dietl P., Spielman W.S., Stanton B.A. Аденозин регулирует хлоридный канал через протеинкиназу C и белок G в клеточной линии кортикального отдела собирательных трубочек кролика. Дж. Клин. расследование 1992;89:834–841. doi: 10.1172/JCI115662. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
83. Sun X.P., Supplisson S., Torres R., Sachs G., Mayer E. Характеристика хлоридных каналов с большой проводимостью в гладких мышцах толстой кишки кролика. Дж. Физиол. 1992; 448: 355–382. doi: 10.1113/jphysiol.1992.sp019046. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
84. Харди С.П., Вальверде М.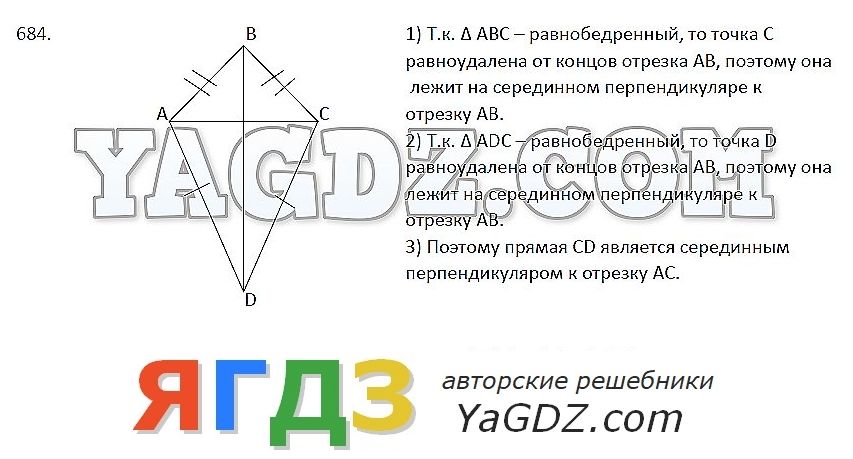 А. Новое действие эстрогенов и антиэстрогенов на плазматическую мембрану, выявленное их регуляцией хлоридного канала с большой проводимостью. ФАСЭБ Дж. 1994;8:760–765. doi: 10.1096/fasebj.8.10.8050676. [PubMed] [CrossRef] [Google Scholar]
А. Новое действие эстрогенов и антиэстрогенов на плазматическую мембрану, выявленное их регуляцией хлоридного канала с большой проводимостью. ФАСЭБ Дж. 1994;8:760–765. doi: 10.1096/fasebj.8.10.8050676. [PubMed] [CrossRef] [Google Scholar]
85. Diaz M., Bahamonde M.I., Lock H., Munoz F.J., Hardy S.P., Posas F., Valverde M.A. Кислоточувствительная активация каналов Maxi Cl(-) Okadaic с помощью трифенилэтиленовые антиэстрогены в клетках нейробластомы мыши С1300. Дж. Физиол. 2001; 536: 79–88. doi: 10.1111/j.1469-7793.2001.00079.x. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
86. Pahapill P.A., Schlichter L.C. Cl(-) каналы в интактных Т-лимфоцитах человека. Дж. Член. биол. 1992;125:171–183. doi: 10.1007/BF00233356. [PubMed] [CrossRef] [Google Scholar]
87. Нам Дж. Х., Чжэн Х. Ф., Эрм К. Х., Ко Дж. Х., Ли И. Дж., Канг Т. М., Ким Т. Дж., Эрм Э., Ким С. Дж. Зависимый от напряжения медленно активирующий анионный ток, регулируемый температурой и внеклеточным рН в В-клетках мыши. Пфлуг. Арка 2006; 452: 707–717. doi: 10.1007/s00424-006-0084-3. [PubMed] [CrossRef] [Google Scholar]
Пфлуг. Арка 2006; 452: 707–717. doi: 10.1007/s00424-006-0084-3. [PubMed] [CrossRef] [Google Scholar]
88. Швиберт Э.М., Лайт Д.Б., Фейес-Тот Г., Нарай-Фейес-Тот А., Стэнтон Б.А. GTP-связывающий белок активирует хлоридные каналы в почечном эпителии. Дж. Биол. хим. 1990;265:7725–7728. doi: 10.1016/S0021-9258(19)38987-2. [PubMed] [CrossRef] [Google Scholar]
89. McGill J.M., Basavappa S., Fitz J.G. Характеристика анионных каналов высокой проводимости в эпителиальных клетках желчных протоков крыс. Являюсь. Дж. Физиол. 1992; 262:G703–G710. doi: 10.1152/jpgi.1992.262.4.G703. [PubMed] [CrossRef] [Google Scholar]
90. McGill J.M., Gettys T.W., Basavappa S., Fitz J.G. GTP-связывающие белки регулируют анионные каналы с высокой проводимостью в эпителиальных клетках желчных протоков крыс. Дж. Член. биол. 1993;133:253–261. doi: 10.1007/BF00232024. [PubMed] [CrossRef] [Google Scholar]
91. Ислам М.Р., Окада Т., Мерзляк П.Г., Тойчиев А.Х., Андо-Акацука Ю., Сабиров Р.З. , Окада Ю. Аннексин А2-С100А10 представляет собой регуляторный компонент Макси- Канал Cl зависит от дефосфорилирования белка тирозина и внутриклеточного Ca 2+ Cell. Физиол. Биохим. 2020; 54: 538–555. doi: 10.33594/000000238. [PubMed] [CrossRef] [Google Scholar]
, Окада Ю. Аннексин А2-С100А10 представляет собой регуляторный компонент Макси- Канал Cl зависит от дефосфорилирования белка тирозина и внутриклеточного Ca 2+ Cell. Физиол. Биохим. 2020; 54: 538–555. doi: 10.33594/000000238. [PubMed] [CrossRef] [Google Scholar]
92. Де Пинто В. Возрождение VDAC: новое понимание семейства белков на границе между митохондриями и цитозолем. Биомолекулы. 2021;11:107. дои: 10.3390/биом11010107. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
93. Розенкранс В.М., Раджендран М., Безруков С.М., Ростовцева Т.К. Регуляция VDAC потока митохондриального кальция: от биофизики канала к болезни. Клеточный кальций. 2021;94:102356. doi: 10.1016/j.ceca.2021.102356. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
94. Коломбини М. Регулирование напряжения в митохондриальном канале, VDAC. Дж. Член. биол. 1989; 111: 103–111. doi: 10.1007/BF01871775. [PubMed] [CrossRef] [Академия Google]
95. Коломбини М. VDAC: Канал на границе между митохондриями и цитозолем.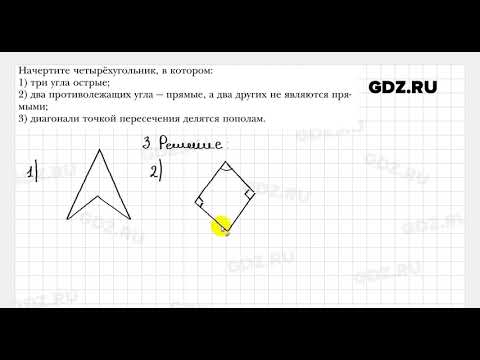 Мол. Клетка. Биохим. 2004; 256–257: 107–115. doi: 10.1023/B:MCBI.0000009862.17396.8d. [PubMed] [CrossRef] [Google Scholar]
Мол. Клетка. Биохим. 2004; 256–257: 107–115. doi: 10.1023/B:MCBI.0000009862.17396.8d. [PubMed] [CrossRef] [Google Scholar]
96. Коломбини М. Стробирование напряжения в VDAC: к молекулярному механизму. В: Миллер С., редактор. Восстановление ионного канала. Пленум Пресс; Нью-Йорк, штат Нью-Йорк, США: 1986. стр. 533–550. [Google Scholar]
97. Шошан-Бармац В., Де Пинто В., Цвекштеттер М., Равив З., Кейнан Н., Арбель Н. VDAC, многофункциональный митохондриальный белок, регулирующий жизнь и смерть клеток. Мол. Асп. Мед. 2010; 31: 227–285. doi: 10.1016/j.mam.2010.03.002. [PubMed] [CrossRef] [Академия Google]
98. Ростовцева Т., Коломбини М. Поток АТФ контролируется потенциалзависимым каналом внешней мембраны митохондрий. Дж. Биол. хим. 1996; 271:28006–28008. doi: 10.1074/jbc.271.45.28006. [PubMed] [CrossRef] [Google Scholar]
99. Ростовцева Т.К., Безруков С.М. Транспорт АТФ по одному митохондриальному каналу, VDAC, изучали с помощью анализа флуктуаций тока. Биофиз. Дж. 1998; 74: 2365–2373. doi: 10.1016/S0006-3495(98)77945-7. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Дж. 1998; 74: 2365–2373. doi: 10.1016/S0006-3495(98)77945-7. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
100. Ростовцева Т.К., Комаров А., Безруков С.М., Коломбини М. Динамика нуклеотидов в каналах VDAC: Генерация структурно-специфического шума. Биофиз. Дж. 2002; 82: 193–205. doi: 10.1016/S0006-3495(02)75386-1. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
101. Dermietzel R., Hwang T.K., Buettner R., Hofer A., Dotzler E., Kremer M., Deutzmann R., Thinnes F.P., Fishman Г.И., Спрей Д.К. и др. Клонирование и локализация in situ порина головного мозга, который образует анионный канал с большой проводимостью в плазматических мембранах астроцитов. проц. Натл. акад. науч. США. 1994;91:499–503. doi: 10.1073/pnas.91.2.499. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
102. Buettner R., Papoutsoglou G., Scemes E., Spray D.C., Dermietzel R. Доказательства локализации секреторного пути изоформы потенциалзависимого анионного канала . проц. Натл. акад. науч. США. 2000;97:3201–3206. doi: 10.1073/pnas.97.7.3201. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
проц. Натл. акад. науч. США. 2000;97:3201–3206. doi: 10.1073/pnas.97.7.3201. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
103. Бахамонде М.И., Вальверде М.А. Потенциал-зависимый анионный канал локализуется в плазматической мембране и периферических, но не перинуклеарных митохондриях. Пфлуг. Арка 2003;446:309–313. doi: 10.1007/s00424-003-1054-7. [PubMed] [CrossRef] [Google Scholar]
104. Бахамонде М.И., Фернандес-Фернандес Дж.М., Гикс Ф.Х., Васкес Э., Вальверде М.А. Потенциал-зависимый анионный канал плазматической мембраны опосредует активируемые антиэстрогенами макси-токи Cl в клетках нейробластомы С1300 . Дж. Биол. хим. 2003; 278:33284–33289. doi: 10.1074/jbc.M302814200. [PubMed] [CrossRef] [Google Scholar]
105. Элиндер Ф., Аканда Н., Тофиги Р., Симидзу С., Цудзимото Ю., Оррениус С., Чеккателли С. Открытие потенциалзависимых анионных каналов плазматической мембраны (VDAC) предшествует активации каспазы при апоптозе нейронов, индуцированном токсическими стимулами. Клеточная смерть. Отличаться. 2005 г.: 10.1038/sj.cdd.4401646. [PubMed] [CrossRef] [Академия Google]
Клеточная смерть. Отличаться. 2005 г.: 10.1038/sj.cdd.4401646. [PubMed] [CrossRef] [Академия Google]
106. Аканда Н., Элиндер Ф. Биофизические свойства потенциал-зависимого анионного канала плазматической мембраны, индуцирующего апоптоз. Биофиз. Дж. 2006; 90:4405–4417. doi: 10.1529/biophysj.105.080028. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
107. Судзуки М., Мизуно А. Новое семейство человеческих каналов Cl(-), связанное с локусом нелетающих дрозофил. Дж. Биол. хим. 2004; 279:22461–22468. doi: 10.1074/jbc.M313813200. [PubMed] [CrossRef] [Google Scholar]
108. Suzuki M. Семейство дрозофил tweety: молекулярные кандидаты на Ca 9 с большой проводимостью0145 2+ -активированные Cl(-) каналы. Эксп. Физиол. 2006; 91: 141–147. doi: 10.1113/expphysiol.2005.031773. [PubMed] [CrossRef] [Google Scholar]
109. Окада Ю., Сато К., Тойчиев А.Х., Судзуки М., Датта А.К., Иноуэ Х., Сабиров Р. Загадки объемно-активируемых анионных каналов. В: Альварес-Лифманс Ф. Дж., Делпир Э., редакторы. Физиология и патология транспортеров хлоридов и каналов в нервной системе. От молекул к болезням. Эльзевир; Сан-Диего, Калифорния, США: 2009. стр. 283–306. [Академия Google]
Дж., Делпир Э., редакторы. Физиология и патология транспортеров хлоридов и каналов в нервной системе. От молекул к болезням. Эльзевир; Сан-Диего, Калифорния, США: 2009. стр. 283–306. [Академия Google]
110. Окада Т., Ислам М.Р., Мерзляк П.Г., Сабиров Р., Окада Ю. Попытки идентификации гена, кодирующего макси-анионный канал. Дж. Физиол. науч. 2012;62:S85. [Google Scholar]
111. Ислам М.Р., Мерзляк П.Г., Окада Т., Сабиров Р.З., Окада Ю. Поиски молекулярной основы максианионного канала. Дж. Физиол. науч. 2013;63:S133. [Google Scholar]
112. Мерзляк П.Г., Окада Ю., Сабиров Р.З. Активность макси-анионного канала в пузырьках мембраны и после восстановления в искусственных мембранах. Дж. Физиол. науч. 2010;60:S123. [Академия Google]
113. Сабиров Р.З., Мерзляк П.Г., Окада Т., Ислам М.Р., Урамото Х., Мори Т., Макино Ю., Мацуура Х., Се Ю., Окада Ю. Переносчик органических анионов SLCO2A1 составляет основной компонент канал Maxi-Cl. EMBO J. 2017; 36:3309–3324. doi: 10.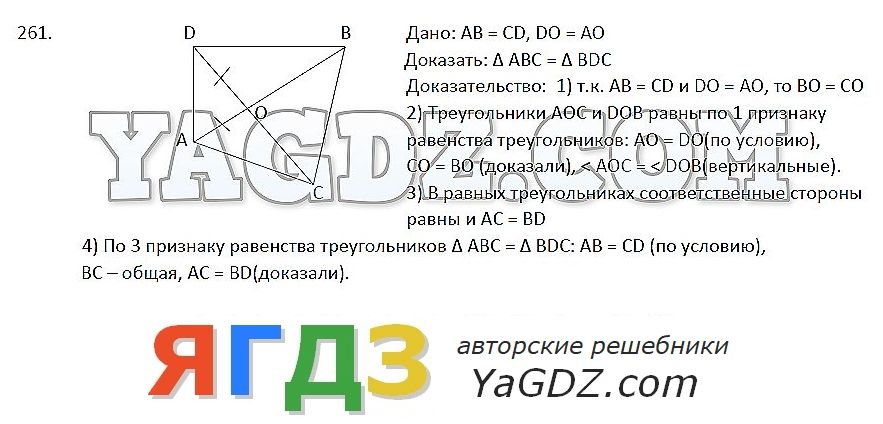 15252/embj.201796685. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
15252/embj.201796685. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
114. Kanai N., Lu R., Satriano J.A., Bao Y., Wolkoff A.W., Schuster V.L. Идентификация и характеристика транспортера простагландинов. Наука. 1995;268:866–869. doi: 10.1126/science.7754369. [PubMed] [CrossRef] [Google Scholar]
115. Чан Б.С., Бао Ю., Шустер В.Л. Роль консервативных трансмембранных катионных аминокислот в переносчике простагландинов PGT. Биохимия. 2002;41:9215–9221. doi: 10.1021/bi0203031. [PubMed] [CrossRef] [Google Scholar]
116. Zhang Z., Xia W., He J., Zhang Z., Ke Y., Yue H., Wang C., Zhang H., Gu J., Ху В. и др. Секвенирование экзома идентифицирует мутации SLCO2A1 как причину первичной гипертрофической остеоартропатии. Являюсь. Дж. Хам. Жене. 2012;90:125–132. doi: 10.1016/j.ajhg.2011.11.019. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
117. Zhang Z., He J.W., Fu W.Z., Zhang C.Q., Zhang Z.L. Две новые мутации в гене SLCO2A1 у китайского пациента с первичной гипертрофической остеоартропатией. Ген. 2014; 534:421–423. doi: 10.1016/j.gene.2013.10.051. [PubMed] [CrossRef] [Google Scholar]
Ген. 2014; 534:421–423. doi: 10.1016/j.gene.2013.10.051. [PubMed] [CrossRef] [Google Scholar]
118. Шустер В.Л. Молекулярные механизмы транспорта простагландинов. Анну. Преподобный Физиол. 1998; 60: 221–242. doi: 10.1146/annurev.physiol.60.1.221. [PubMed] [CrossRef] [Академия Google]
119. Шустер В.Л. Транспорт простагландинов. Простагландины Другие липиды Медиат. 2002; 68–69: 633–647. doi: 10.1016/S0090-6980(02)00061-8. [PubMed] [CrossRef] [Google Scholar]
120. Chang H.Y., Locker J., Lu R., Schuster V.L. Неспособность закрыть постнатальный артериальный проток у мышей с дефицитом транспортера простагландина. Тираж. 2010;121:529–536. doi: 10.1161/CIRCULATIONAHA.109.862946. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
121. Шустер В.Л., Чи Ю., Лу Р. Транспортер простагландинов: обратный захват эйкозаноидов, контроль передачи сигналов и разработка высокоаффинных ингибиторов в качестве кандидатов в лекарства . Транс. Являюсь. клин. Клим. доц. 2015; 126: 248–257. [Бесплатная статья PMC] [PubMed] [Google Scholar]
2015; 126: 248–257. [Бесплатная статья PMC] [PubMed] [Google Scholar]
122. Майнор Д.Л., младший. Перелистывая каналы, обнаруживает транспортер двойного назначения. EMBO J. 2017; 36:3272–3273. doi: 10.15252/embj.201798304. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
123. Picollo A., Malvezzi M., Accardi A. Белки TMEM16: неизвестная структура и запутанные функции. Дж. Мол. биол. 2015; 427:94–105. doi: 10.1016/j.jmb.2014.09.028. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
124. Пиколло А., Пуш М. Хлоридная/протонная антипортная активность белков CLC млекопитающих ClC-4 и ClC-5. Природа. 2005; 436:420–423. doi: 10.1038/nature03720. [PubMed] [CrossRef] [Академия Google]
125. Шеел О., Здебик А.А., Лурдель С., Йенч Т.Дж. Зависимый от напряжения электрогенный хлоридный/протонный обмен эндосомальными белками CLC. Природа. 2005; 436: 424–427. doi: 10.1038/nature03860. [PubMed] [CrossRef] [Google Scholar]
126. Accardi A. , Picollo A. CLC-каналы и транспортеры: белки с пограничными особенностями. Биохим. Биофиз. Акта. 2010;1798:1457–1464. doi: 10.1016/j.bbamem.2010.02.022. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
, Picollo A. CLC-каналы и транспортеры: белки с пограничными особенностями. Биохим. Биофиз. Акта. 2010;1798:1457–1464. doi: 10.1016/j.bbamem.2010.02.022. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
127. Fahlke C., Kortzak D., Machtens J.P. Молекулярная физиология анионных каналов EAAT. Пфлуг. Арка 2016;468:491–502. doi: 10.1007/s00424-015-1768-3. [PubMed] [CrossRef] [Google Scholar]
128. Охана Э., Ян Д., Щейников Н., Муаллем С. Различные способы переноса переносчиков растворенных веществ 26 семейства переносчиков анионов. Дж. Физиол. 2009; 587: 2179–2185. doi: 10.1113/jphysiol.2008.164863. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
129. Riquelme G., Llanos P., Tischner E., Neil J., Campos B. Аннексин 6 модулирует максихлоридный канал апикальной мембраны синцитиотрофобласта, выделенного из плаценты человека. Дж. Биол. хим. 2004;279: 50601–50608. doi: 10.1074/jbc.M407859200. [PubMed] [CrossRef] [Google Scholar]
130. Santamaria-Kisiel L. , Rintala-Dempsey AC, Shaw GS. Кальций-зависимые и -независимые взаимодействия белков семейства S100. Биохим. Дж. 2006; 396: 201–214. doi: 10.1042/BJ20060195. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
, Rintala-Dempsey AC, Shaw GS. Кальций-зависимые и -независимые взаимодействия белков семейства S100. Биохим. Дж. 2006; 396: 201–214. doi: 10.1042/BJ20060195. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
131. Пауэлл М.А., Гленни Дж.Р. Регуляция связывания фосфолипидов кальпактина I путем связывания легкой цепи кальпактина I и фосфорилирования с помощью p60v-src. Биохим. Дж. 1987;247:321–328. doi: 10.1042/bj2470321. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
132. Нилиус Б., Герке В., Пренен Дж., Шукс Г., Хейнке С., Вебер К., Другманс Г. Аннексин II модулирует громкость -активация хлоридных токов в эндотелиальных клетках сосудов. Дж. Биол. хим. 1996; 271:30631–30636. doi: 10.1074/jbc.271.48.30631. [PubMed] [CrossRef] [Google Scholar]
133. Borthwick L.A., McGaw J., Conner G., Taylor C.J., Gerke V., Mehta A., Robson L., Muimo R. Образование цАМФ/белка Зависимый от киназы А комплекс аннексина 2-S100A10 с белком-регулятором проводимости при муковисцидозе (CFTR) регулирует функцию канала CFTR. Мол. биол. Клетка. 2007; 18: 3388–339.7. doi: 10.1091/mbc.e07-02-0126. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Мол. биол. Клетка. 2007; 18: 3388–339.7. doi: 10.1091/mbc.e07-02-0126. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
134. Borthwick L.A., Riemen C., Goddard C., Colledge WH, Mehta A., Gerke V., Muimo R. Дефектное формирование PKA/CnA Комплекс -зависимый аннексин 2-S100A10/CFTR в клетках муковисцидоза deltaF508. Клетка. Сигнал. 2008;20:1073–1083. doi: 10.1016/j.cellsig.2008.01.021. [PubMed] [CrossRef] [Google Scholar]
135. Muimo R. Регуляция функции CFTR комплексом аннексина A2-S100A10 в норме и при патологии. Ген. физиол. Биофиз. 2009 г.;28:F14–F19. [PubMed] [Google Scholar]
136. Cahalan MD, Lewis R.S. Роль калиевых и хлоридных каналов в регуляции объема Т-лимфоцитами. соц. Ген. физиол. сер. 1988; 43: 281–301. [PubMed] [Google Scholar]
137. Schlichter LC, Grygorczyk R., Pahapill P.A., Grygorczyk C. Большой хлоридный канал с множественной проводимостью в нормальных Т-лимфоцитах человека. Пфлуг. Арка 1990; 416:413–421. doi: 10.1007/BF00370748. [PubMed] [CrossRef] [Google Scholar]
[PubMed] [CrossRef] [Google Scholar]
138. Hurnak O., Zachar J. Максихлоридные каналы в миобластах L6. Ген. физиол. Биофиз. 1992;11:389–400. [PubMed] [Google Scholar]
139. Митчелл Ч.Х., Ван Л., Джейкоб Т.Дж. Хлоридный канал с большой проводимостью в клетках пигментированного цилиарного эпителия, активированный GTPgammaS. Дж. Член. биол. 1997; 158: 167–175. doi: 10.1007/s002329
4. [PubMed] [CrossRef] [Google Scholar]140. Bernucci L., Umana F., Llanos P., Riquelme G. Большой хлоридный канал из преэклампсии плаценты человека. Плацента. 2003; 24:895–903. doi: 10.1016/S0143-4004(03)00144-9. [PubMed] [CrossRef] [Академия Google]
141. МакЛарнон Дж.Г., Ким С.У. Ионные каналы в культивируемых шванновских клетках взрослого человека. Глия. 1991; 4: 534–539. doi: 10.1002/glia.440040513. [PubMed] [CrossRef] [Google Scholar]
142. Quasthoff S., Strupp M., Grafe P. Анионный канал с высокой проводимостью в везикулах шванновских клеток из спинномозговых корешков крысы. Глия. 1992; 5:17–24. doi: 10.1002/glia.440050104. [PubMed] [CrossRef] [Google Scholar]
Глия. 1992; 5:17–24. doi: 10.1002/glia.440050104. [PubMed] [CrossRef] [Google Scholar]
143. Томпсон Р.Дж., Нордин М.Х., Хауэлл К.Е., Колдуэлл Дж.Х. Анионный канал комплекса Гольджи с большой проводимостью. Биофиз. Дж. 2002; 83: 278–289.. doi: 10.1016/S0006-3495(02)75168-0. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
144. Do C.W., Peterson-Yantorno K., Mitchell C.H., Civan M.M. цАМФ-активированные макси-Cl (-) каналы в нативных бычьих пигментированных цилиарных эпителиальных клетках. Являюсь. Дж. Физиол. Клеточная физиол. 2004; 287:C1003–C1011. doi: 10.1152/ajpcell.00175.2004. [PubMed] [CrossRef] [Google Scholar]
145. Do CW, Civan M.M. Активированные набуханием хлоридные каналы в образовании водянистой влаги: с одной и с другой стороны. Акта Физиол. 2006; 187: 345–352. doi: 10.1111/j.1748-1716.2006.01548.x. [PubMed] [CrossRef] [Академия Google]
146. Becq F., Fanjul M., Mahieu I., Berger Z., Gola M., Hollande E. Анионные каналы в клеточной линии рака поджелудочной железы человека (Capan-1) протокового происхождения. Пфлуг. Арка 1992; 420:46–53. doi: 10.1007/BF00378640. [PubMed] [CrossRef] [Google Scholar]
Пфлуг. Арка 1992; 420:46–53. doi: 10.1007/BF00378640. [PubMed] [CrossRef] [Google Scholar]
147. Schneider G.T., Cook D.I., Gage P.W., Young J.A. Чувствительные к напряжению хлоридные каналы с высокой проводимостью в просветной мембране культивируемых легочных альвеолярных (тип II) клеток. Пфлуг. Арка 1985; 404: 354–357. doi: 10.1007/BF00585348. [PubMed] [CrossRef] [Академия Google]
148. Vallejos C., Riquelme G. Максихлоридный канал в синцитиотрофобласте человека: путь оттока таурина в регуляции плацентарного объема? Плацента. 2007; 28:1182–1191. doi: 10.1016/j.placenta.2007.06.005. [PubMed] [CrossRef] [Google Scholar]
149. Стеа А., медсестра К.А. Хлоридные каналы в культивируемых гломусных клетках каротидного тела крысы. Являюсь. Дж. Физиол. 1989; 257:C174–C181. doi: 10.1152/ajpcell.1989.257.2.C174. [PubMed] [CrossRef] [Google Scholar]
150. Хах Б.С., Северный Р.А. Нейромодуляция внеклеточными рецепторами АТФ и Р2Х в ЦНС. Нейрон. 2012; 76: 51–69. doi: 10. 1016/j.neuron.2012.09.024. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
1016/j.neuron.2012.09.024. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
151. Бернсток Г., Пеллег А. Сердечная пуринергическая сигнализация в норме и при заболеваниях. Пуринергический сигнал. 2015; 11:1–46. doi: 10.1007/s11302-014-9436-1. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
152. Лю Х.Т., Тойчиев А.Х., Такахаши Н., Сабиров Р.З., Окада Ю. Макси-анионный канал как кандидатный путь высвобождения осмочувствительного АТФ из астроцитов мыши в первичной культуре. Сотовый рез. 2008 г.: 10.1038/cr.2008.49. [PubMed] [CrossRef] [Google Scholar]
153. Matsuura H., Kojima A., Fukushima Y., Xie Y., Mi X., Sabirov R.Z., Okada Y. Положительные инотропные эффекты АТФ, высвобождаемой через макси- анионный канал в перфузированных по Лангендорфу сердцах мышей, подвергшихся ишемии-реперфузии. Фронт. Сотовый Дев. биол. 2021;9:597997. doi: 10.3389/fcell.2021.597997. [Статья PMC free] [PubMed] [CrossRef] [Google Scholar]
154. Терновский В.И., Окада Ю. , Сабиров Р.З. Определение размера пор объемно-чувствительного анионного канала путем дифференциального разделения полимера. ФЭБС лат. 2004; 576: 433–436. doi: 10.1016/j.febslet.2004.090,051. [PubMed] [CrossRef] [Google Scholar]
, Сабиров Р.З. Определение размера пор объемно-чувствительного анионного канала путем дифференциального разделения полимера. ФЭБС лат. 2004; 576: 433–436. doi: 10.1016/j.febslet.2004.090,051. [PubMed] [CrossRef] [Google Scholar]
155. Nakanishi T., Nakamura Y., Umeno J. Последние достижения в исследованиях SLCO2A1 как ключевого регулятора доставки простагландинов к местам их действия. Фармакол. тер. 2021;223:107803. doi: 10.1016/j.pharmthera.2021.107803. [PubMed] [CrossRef] [Google Scholar]
156. Умено Дж., Хисамацу Т., Эсаки М., Хирано А., Кубокура Н., Асано К., Кочи С., Янаи С., Фуюно Ю., Шимамура К. и др. Наследственная энтеропатия, вызванная мутациями в гене SLCO2A1, кодирующем переносчик простагландинов. Генетика PLoS. 2015;11:e1005581. doi: 10.1371/journal.pgen.1005581. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
157. Хосоэ Н., Омия Н., Хираи Ф., Умено Дж., Эсаки М., Ямагами Х., Онодера К., Бамба С., Имаэда Х., Янаи С. и др. Хроническая энтеропатия, связанная с геном SLCO2A1 [CEAS] — характеристика кишечного расстройства, которую следует учитывать при дифференциальной диагностике болезни Крона. Дж. Кронс. Колит. 2017;11:1277–1281. doi: 10.1093/ecco-jcc/jjx068. [PubMed] [CrossRef] [Google Scholar]
Дж. Кронс. Колит. 2017;11:1277–1281. doi: 10.1093/ecco-jcc/jjx068. [PubMed] [CrossRef] [Google Scholar]
158. Цузуки Ю., Аояги Р., Миягути К., Ашитани К., Ого Х., Ямаока М., Исидзава К., Каяно Х., Хисамацу Т., Умено Дж. и др. Хроническая энтеропатия, ассоциированная с SLCO2A1, с пахидермопериостозом. Стажер Мед. 2020;59: 3147–3154. doi: 10.2169/internalmedicine.4756-20. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
159. Sonoda A., Wada Y., Togo K., Mizukami K., Fuyuno Y., Umeno J., Fujioka S., Fukuda K. , Окамото К., Огава Р. и др. Характерный внешний вид лица был ключом к диагностике хронической энтеропатии, связанной с первичной гипертрофической остеоартропатией, связанной с SLCO2A1. Стажер Мед. 2020; 59: 491–494. doi: 10.2169/internalmedicine.3369-19. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
160. Guda K., Fink S.P., Milne G.L., Molyneaux N., Ravi L., Lewis S.M., Dannenberg A.J., Montgomery C.G., Zhang S., Willis J., et al. Инактивирующая мутация в гене транспортера простагландина, SLCO2A1, связана с семейной группировкой пальцев, неоплазией толстой кишки и устойчивостью к НПВП. Рак Пред. Рез. 2014;7:805–812. doi: 10.1158/1940-6207.CAPR-14-0108. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Инактивирующая мутация в гене транспортера простагландина, SLCO2A1, связана с семейной группировкой пальцев, неоплазией толстой кишки и устойчивостью к НПВП. Рак Пред. Рез. 2014;7:805–812. doi: 10.1158/1940-6207.CAPR-14-0108. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
161. Джимбо К., Окуно Т., Огаки Р., Нишикубо К., Китамура Ю., Сакураи Ю., Куан Л., Сёдзи Х. , Канаи Ю., Симидзу Т. и др. Новая мутация в гене SLCO2A1, кодирующем переносчик простагландинов, вызывает хроническую энтеропатию. ПЛОС ОДИН. 2020;15:e0241869. doi: 10.1371/journal.pone.0241869. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
162. Наканиши Т., Оно Ю., Аотани Р., Маруяма С., Шимада Х., Камо С., Осима Х., Осима М. , Schuetz JD, Tamai I. Новая роль OATP2A1/SLCO2A1 в мышиной модели рака толстой кишки. науч. 2017;7:16567. doi: 10.1038/s41598-017-16738-y. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
163. Lopes C., Pereira C., Farinha M., Medeiros R., Dinis-Ribeiro M. Путь простагландина E2 нарушен при аденокарциноме желудка в Кавказское население. Междунар. Дж. Мол. науч. 2020;21:7680. дои: 10.3390/ijms21207680. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Путь простагландина E2 нарушен при аденокарциноме желудка в Кавказское население. Междунар. Дж. Мол. науч. 2020;21:7680. дои: 10.3390/ijms21207680. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
164. Ван Ю., Ма С., Руццо В.Л. Пространственное моделирование экспрессии метаболических генов рака предстательной железы выявляет обширную гетерогенность и избирательную уязвимость. науч. Респ. 2020; 10:3490. doi: 10.1038/s41598-020-60384-w. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
165. Tang L., Zhu Q., Wang Z., Shanahan C.M., Bensen J.T., Fontham E.T.H., Smith G.J., Pop E.A., Azabdaftari G., Молер Дж.Л. и соавт. Дифференциальные ассоциации транспортеров SLCO с агрессивностью рака предстательной железы у афроамериканцев и американцев европейского происхождения. Эпидемиол рака. Биомарк. Пред. 2021 г.: 10.1158/1055-9965.ЭПИ-20-1389. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
166. Cai C., Long Y., Li Y., Huang M. Сосуществование COX7A2L-ALK, LINC01210-ALK, ATP13A4-ALK и приобретенного SLCO2A1 -ALK при аденокарциноме легкого с потерей перестроек при лечении кризотинибом и церитинибом: клинический случай. Онко. Цели Тер. 2020;13:8313–8316. doi: 10.2147/OTT.S258067. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Онко. Цели Тер. 2020;13:8313–8316. doi: 10.2147/OTT.S258067. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
167. Наката Р., Накамура Ю., Хосоми С., Окуда Х., Нисида Ю., Сугита Н., Итани С., Надатани Ю. , Отани К., Танака Ф. и др. Дефицит Slco2a1 усугубляет экспериментальный колит за счет активации воспаления в макрофагах: возможный механизм хронической энтеропатии, связанный с геном SLCO2A1. науч. Респ. 2020; 10:4883. дои: 10.1038/s41598-020-61775-9. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
168. Инагаки М., Нисимура Т., Наканиши Т., Шимада Х., Ногучи С., Аканума С.И., Тачикава М., Хосоя К.И., Тамаи И., Накашима Э. и др. Вклад транспортера простагландинов OATP2A1/SLCO2A1 в передачу сигналов гормонов от плаценты к матери и индукцию родов. iНаука. 2020;23:101098. doi: 10.1016/j.isci.2020.101098. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
169. Наканиши Т., Сакияма С., Такашима Х., Хонда Р., Шумба М.Н., Накамура Ю. , Касахара К., Тамаи И. Токсикологические значение ингибирования переносчика простагландинов SLCO2A1 сигаретным дымом при обострении воспаления легких. Токсикол. заявл. Фармакол. 2020;405:115201. doi: 10.1016/j.taap.2020.115201. [PubMed] [CrossRef] [Академия Google]
, Касахара К., Тамаи И. Токсикологические значение ингибирования переносчика простагландинов SLCO2A1 сигаретным дымом при обострении воспаления легких. Токсикол. заявл. Фармакол. 2020;405:115201. doi: 10.1016/j.taap.2020.115201. [PubMed] [CrossRef] [Академия Google]
170. Норита К., Асанума К., Койке Т., Оката Т., Фудзия Т., Абэ Ю., Накагава К., Хатта В., Уно К., Накамура Т. и др. Нарушение целостности слизистой оболочки проксимального отдела пищевода способствует развитию рефрактерной к ингибиторам протонной помпы неэрозивной рефлюксной болезни. пищеварение. 2020: 1–11. doi: 10.1159/000508661. [PubMed] [CrossRef] [Google Scholar]
171. Theocharidis G., Baltzis D., Roustit M., Tellechea A., Dangwal S., Khetani R.S., Shu B., Zhao W., Fu J., Bhasin С. и др. Интегрированная транскриптомика кожи и мультиплексные анализы сыворотки выявляют новые механизмы заживления ран при диабетических язвах стопы. Диабет. 2020;69: 2157–2169. doi: 10.2337/db20-0188. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Широкополосная зелень
ВП широкополосной зелени являются одними из самых простых показателей общего количества и силы зеленой растительности. Они представляют собой комбинации измерений отражательной способности, которые чувствительны к комбинированным эффектам концентрации хлорофилла в листве, площади листового полога, сгущению листвы и архитектуре полога. Эти VI предназначены для измерения общего количества и качества фотосинтетического материала в растительности, что необходимо для понимания состояния растительности для любых целей. Эти VI являются интегративным измерением этих факторов и хорошо коррелируют с фракционным поглощением фотосинтетически активной радиации (fAPAR) навесами растений и растительными пикселями. Они не предоставляют количественную информацию о каком-либо одном биологическом факторе или факторе окружающей среды, влияющем на fAPAR, но была обнаружена широкая корреляция между широкополосными VI зеленого цвета и LAI растительного покрова.
Они представляют собой комбинации измерений отражательной способности, которые чувствительны к комбинированным эффектам концентрации хлорофилла в листве, площади листового полога, сгущению листвы и архитектуре полога. Эти VI предназначены для измерения общего количества и качества фотосинтетического материала в растительности, что необходимо для понимания состояния растительности для любых целей. Эти VI являются интегративным измерением этих факторов и хорошо коррелируют с фракционным поглощением фотосинтетически активной радиации (fAPAR) навесами растений и растительными пикселями. Они не предоставляют количественную информацию о каком-либо одном биологическом факторе или факторе окружающей среды, влияющем на fAPAR, но была обнаружена широкая корреляция между широкополосными VI зеленого цвета и LAI растительного покрова.
Широкополосные индикаторы зелени сравнивают измерения коэффициента отражения от пика отражения растительности в ближнем инфракрасном диапазоне с другим измерением, проведенным в красном диапазоне, где хлорофилл поглощает фотоны для накопления энергии посредством фотосинтеза. Использование измерений в ближнем инфракрасном диапазоне с гораздо большей глубиной проникновения через полог, чем в красном, позволяет зондировать общее количество зеленой растительности в столбце до тех пор, пока сигнал не достигнет насыщения на очень высоких уровнях. Поскольку спектр этих характеристик довольно широк, многие широкополосные индексы зелени могут эффективно работать даже с данными изображений, полученными с помощью широкополосных мультиспектральных датчиков, таких как AVHRR, Landsat TM и QuickBird. Приложения включают исследования фенологии (роста) растительности, оценки землепользования и климатологического воздействия, а также моделирование продуктивности растительности.
Использование измерений в ближнем инфракрасном диапазоне с гораздо большей глубиной проникновения через полог, чем в красном, позволяет зондировать общее количество зеленой растительности в столбце до тех пор, пока сигнал не достигнет насыщения на очень высоких уровнях. Поскольку спектр этих характеристик довольно широк, многие широкополосные индексы зелени могут эффективно работать даже с данными изображений, полученными с помощью широкополосных мультиспектральных датчиков, таких как AVHRR, Landsat TM и QuickBird. Приложения включают исследования фенологии (роста) растительности, оценки землепользования и климатологического воздействия, а также моделирование продуктивности растительности.
Увеличение концентрации хлорофилла в листьях или площади листа, уменьшение комков листвы и изменения в архитектуре кроны могут способствовать уменьшению длин волн в ближнем ИК-диапазоне и увеличению длин волн в красной области, тем самым вызывая увеличение значений широкополосной зелени.
- Вегетационный индекс разницы (DVI)
- Расширенный вегетационный индекс (EVI)
- Глобальный индекс мониторинга окружающей среды (GEMI)
- Зеленый индекс атмосферостойкости (GARI)
- Индекс зеленого хлорофилла (GCI)
- Зеленый индекс растительности (GDVI)
- Индекс зеленого листа (GLI)
- Зеленый нормализованный разностный индекс вегетации (GNDVI)
- Экологически оптимизированный индекс растительности с поправкой на почву (GOSAVI)
- Зеленое соотношение растительности (GRVI)
- Индекс растительности с поправкой на зеленую почву (GSAVI)
- Индекс зеленой растительности (GVI)
- Инфракрасный процентный индекс растительности (IPVI)
- Индекс площади листьев (LAI)
- Модифицированный нелинейный индекс (MNLI)
- Модифицированное простое отношение (MSR)
- Модифицированный индекс растительности 2 с поправкой на почву (MSAVI2)
- Нелинейный индекс (NLI)
- Нормализованный разностный вегетативный индекс (NDVI)
- Оптимизированный индекс растительности с поправкой на почву (OSAVI)
- Перенормированный разностный индекс вегетации (RDVI)
- Индекс растительности с поправкой на почву (SAVI)
- Простое соотношение (SR)
- Суммированный индекс зеленого (SGI)
- Преобразованный индекс различий растительности (TDVI)
- Треугольный индекс зелени (TGI)
- Индекс стойкости к видимой атмосфере (VARI)
- Индекс растительности с широким динамическим диапазоном (WDRVI)
- Улучшенный индекс растительности WorldView (WV-VI)
Индекс различия растительности (DVI)
Этот индекс различает почву и растительность, но не учитывает разницу между коэффициентом отражения и излучением, вызванную атмосферными эффектами или тенями.
Ссылка: Tucker, C. «Красные и фотографические инфракрасные линейные комбинации для мониторинга растительности. Дистанционное зондирование окружающей среды 8 (1979): 127–150.
Расширенный индекс растительности (EVI)
6 Первоначально разработанный для использования с данными MODIS в качестве улучшения по сравнению с NDVI за счет оптимизации сигнала растительности в областях с высоким индексом площади листьев (LAI). Он наиболее полезен в регионах с высоким LAI, где NDVI может насыщаться. Он использует область синего отражения для коррекции фоновые сигналы почвы и уменьшить влияние атмосферы, включая аэрозольное рассеивание9.0003
Значения EVI должны находиться в диапазоне от 0 до 1 для пикселей растительности. Яркие объекты, такие как облака и белые здания, а также темные объекты, такие как вода, могут привести к аномальным значениям пикселей на изображении EVI. Перед созданием изображения EVI вы должны замаскировать облака и яркие элементы из изображения отражательной способности и, при необходимости, пороговые значения пикселей от 0 до 1. См. раздел «Предварительная обработка» темы Spectral Indices .
Перед созданием изображения EVI вы должны замаскировать облака и яркие элементы из изображения отражательной способности и, при необходимости, пороговые значения пикселей от 0 до 1. См. раздел «Предварительная обработка» темы Spectral Indices .
Ссылка: Huete, A., et al. «Обзор радиометрических и биофизических характеристик индексов растительности MODIS». Дистанционное зондирование окружающей среды 83 (2002): 195–213.
Глобальный индекс мониторинга окружающей среды (GEMI)
Этот нелинейный вегетационный индекс используется для глобального мониторинга окружающей среды на основе спутниковых изображений и пытается скорректировать влияние атмосферы. Он похож на NDVI, но менее чувствителен к атмосферным воздействиям. Поражается голой почвой; поэтому его не рекомендуется использовать на участках с редкой или умеренно густой растительностью.
Где:
Ссылка: Пинти, Б. и М. Верстраете. GEMI: нелинейный индекс для мониторинга глобальной растительности со спутников. Растительность 101 (1992): 15-20.
и М. Верстраете. GEMI: нелинейный индекс для мониторинга глобальной растительности со спутников. Растительность 101 (1992): 15-20.
Зеленый индекс атмосферостойкости (GARI)
Этот индекс более чувствителен к широкому диапазону концентраций хлорофилла и менее чувствителен к атмосферным воздействиям, чем NDVI.
Гамма-постоянная — это весовая функция, которая зависит от аэрозольных условий в атмосфере. ENVI использует значение 1,7, рекомендованное Гительсоном, Кауфманом и Мерзилаком (1996, стр. 296).
Ссылка: Гительсон А., Ю. Кауфман и М. Мерзилак. «Использование зеленого канала для дистанционного зондирования глобальной растительности с помощью EOS-MODIS». Дистанционное зондирование окружающей среды 58 (1996): 289-298.
Индекс зеленого хлорофилла (GCI)
Этот показатель используется для оценки содержания хлорофилла в листьях различных видов растений.
Широкие длины волн в ближнем ИК-диапазоне и зеленом спектре обеспечивают более точное предсказание содержания хлорофилла, обеспечивая при этом большую чувствительность и более высокое отношение сигнал/шум.
Ссылка: Гительсон А., Гриц Ю., Мерзляк М. «Взаимосвязь между содержанием хлорофилла в листьях и спектральной отражательной способностью и алгоритмы неразрушающей оценки хлорофилла в листьях высших растений». Журнал физиологии растений 160 (2003): 271-282.
Вегетационный индекс Green Difference (GDVI)
Первоначально этот индекс был разработан с использованием цветной инфракрасной фотографии для прогнозирования потребности кукурузы в азоте.
Ссылка: Шрипада Р. и др. «Определение сезонных требований к азоту для кукурузы с использованием аэрофотосъемки в цветном инфракрасном диапазоне».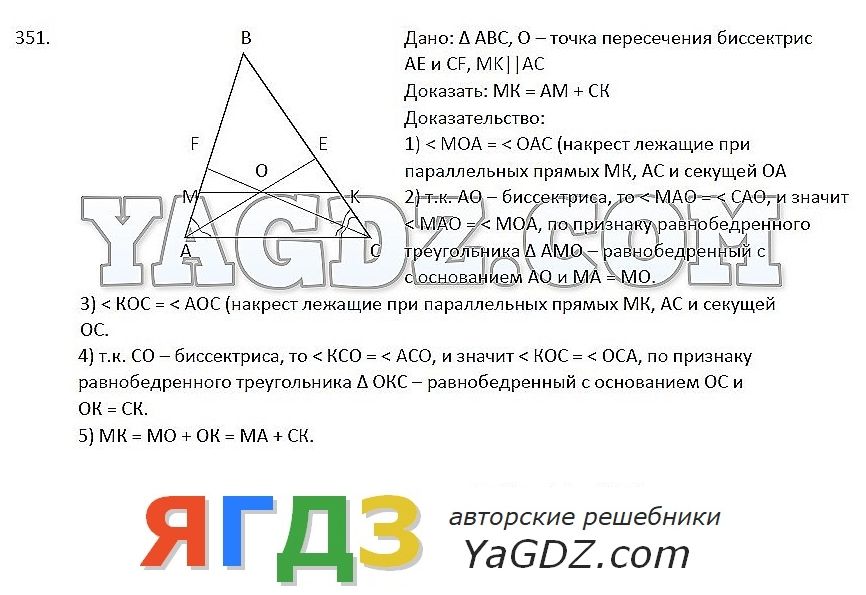 Кандидат наук. диссертация, Государственный университет Северной Каролины, 2005 г.
Кандидат наук. диссертация, Государственный университет Северной Каролины, 2005 г.
Индекс зеленых листьев (GLI)
Этот индекс изначально был разработан для использования с цифровой камерой RGB для измерения покрытия пшеницы, где красные, зеленые и синие цифровые числа (DN) находятся в диапазоне от 0 до 255.
Диапазон значений GLI от -1 до +1. Отрицательные значения представляют почву и неживые элементы, а положительные значения представляют зеленые листья и стебли.
Ссылка: Louhaichi, M., M. Borman, and D. Johnson. «Пространственно расположенная платформа и аэрофотосъемка для документирования воздействия выпаса на пшеницу». Geocarto International 16, № 1 (2001): 65-70.
Зеленый нормализованный разностный индекс растительности (GNDVI)
Этот индекс аналогичен NDVI, за исключением того, что он измеряет зеленый спектр от 540 до 570 нм вместо красного спектра. Этот индекс более чувствителен к концентрации хлорофилла, чем NDVI.
Ссылка: Гительсон А., Мерзляк М. «Дистанционное определение концентрации хлорофилла в листьях высших растений». Достижения в области космических исследований 22 (1998): 689-692.
Индекс растительности с поправкой на зеленый оптимизированный грунт (GOSAVI)
Первоначально этот индекс был разработан с использованием цветной инфракрасной фотографии для прогнозирования потребности кукурузы в азоте. Он похож на OSAVI, но заменяет красную полосу зеленой.
Ссылка: Шрипада Р. и др. «Определение сезонных требований к азоту для кукурузы с использованием аэрофотосъемки в цветном инфракрасном диапазоне». Кандидат наук. диссертация, Государственный университет Северной Каролины, 2005 г.
Индекс растительности зеленого цвета (GRVI)
Этот индекс чувствителен к скорости фотосинтеза в пологе леса, поскольку на коэффициент отражения зеленого и красного сильно влияют изменения в пигментах листьев.
Ссылка: Шрипада Р. и др. «Аэрофотосъемка в цветном инфракрасном диапазоне для определения потребности кукурузы в азоте в начале сезона». Агрономический журнал 98 (2006): 968-977.
Зеленая почва Скорректированный индекс растительности (GSAVI)
Первоначально этот индекс был разработан с использованием цветной инфракрасной фотографии для прогнозирования потребности кукурузы в азоте. Он похож на SAVI, но заменяет красную полосу зеленой.
Ссылка: Шрипада Р. и др. «Определение сезонных требований к азоту для кукурузы с использованием аэрофотосъемки в цветном инфракрасном диапазоне». Кандидат наук. диссертация, Университет штата Северная Каролина, 2005 г.
Индекс зеленой растительности (GVI)
Этот индекс сводит к минимуму влияние фоновой почвы, подчеркивая при этом зеленую растительность. Он использует глобальные коэффициенты, которые взвешивают значения пикселей для создания новых преобразованных полос. Он также известен как индекс зеленой растительности Landsat TM Tasseled Cap. Значения варьируются от -1 до 1.
Он использует глобальные коэффициенты, которые взвешивают значения пикселей для создания новых преобразованных полос. Он также известен как индекс зеленой растительности Landsat TM Tasseled Cap. Значения варьируются от -1 до 1.
GVI изначально был разработан для использования с Landsat TM, но он также будет работать с соответствующими диапазонами Landsat ETM+ и Landsat 8.
Ссылка: Каут Р. и Г. Томас. «Шапка с кисточками — графическое описание спектрально-временного развития сельскохозяйственных культур с точки зрения Landsat» в Материалы симпозиума LARS 1976 г. по машинной обработке данных дистанционного зондирования , Уэст-Лафайет, Индиана: Университет Пердью, стр. 4B41-4B51.
Инфракрасный процентный индекс растительности (IPVI)
Этот индекс функционально аналогичен NDVI, но в вычислительном отношении он быстрее. Диапазон значений от 0 до 1.
Диапазон значений от 0 до 1.
Ссылка: Криппен, Р. «Ускоренный расчет вегетационного индекса». Дистанционное зондирование окружающей среды 34 (1990): 71-73.
Индекс площади листьев (LAI)
Этот индекс используется для оценки листового покрова и прогнозирования роста и урожайности культур. ENVI рассчитывает зеленый LAI, используя следующую эмпирическую формулу из Boegh et al (2002):
Где EVI — значение Расширенного индекса растительности.
Высокие значения LAI обычно находятся в диапазоне примерно от 0 до 3,5. Однако, когда сцена содержит облака и другие яркие элементы, создающие насыщенные пиксели, значения LAI могут превышать 3,5. Вы должны в идеале замаскировать облака и яркие детали из вашей сцены перед созданием изображения LAI.
Ссылка: Boegh, E., H. Soegaard, N. Broge, C. Hasager, N. Jensen, K. Schelde и A. Thomsen. «Мультиспектральные данные по воздуху для количественной оценки индекса площади листьев, концентрации азота и эффективности фотосинтеза в сельском хозяйстве». Дистанционное зондирование окружающей среды 81, №. 2-3 (2002): 179-193.
Thomsen. «Мультиспектральные данные по воздуху для количественной оценки индекса площади листьев, концентрации азота и эффективности фотосинтеза в сельском хозяйстве». Дистанционное зондирование окружающей среды 81, №. 2-3 (2002): 179-193.
Модифицированный нелинейный индекс (MNLI)
Этот индекс является усовершенствованием нелинейного индекса (NLI), который включает скорректированный почвенный индекс растительности (SAVI) для учета фона почвы. ENVI использует коэффициент корректировки фона купола ( L ) значение 0,5.
Ссылка: Ян, З., П. Уиллис и Р. Мюллер. «Влияние изображения AWIFS с улучшенным соотношением полос на точность классификации культур». Proceedings of the Pecora 17 Remote Sensing Symposium (2008), Денвер, Колорадо.
Модифицированный индекс растительности с поправкой на почву 2 (MSAVI2)
Этот индекс является упрощенной версией индекса MSAVI, предложенного Qi, et al (1994). ), который улучшает индекс растительности с поправкой на почву (SAVI). Это уменьшает шум почвы и увеличивает динамический диапазон сигнала растительности. MSAVI2 основан на индуктивном методе, в котором не используется константа 9.0604 L Значение (как в SAVI), чтобы выделить здоровую растительность.
), который улучшает индекс растительности с поправкой на почву (SAVI). Это уменьшает шум почвы и увеличивает динамический диапазон сигнала растительности. MSAVI2 основан на индуктивном методе, в котором не используется константа 9.0604 L Значение (как в SAVI), чтобы выделить здоровую растительность.
Ссылка: Qi, J., A. Chehbouni, A. Huete, Y. Kerr, and S. Soroushian. «Модифицированный индекс растительности с поправкой на почву». Дистанционное зондирование окружающей среды 48 (1994): 119-126.
Модифицированный простой коэффициент (MSR)
Этот индекс был разработан в качестве улучшения по сравнению с RDVI путем объединения в формуле простого коэффициента. В результате повышается чувствительность к биофизическим параметрам растительности.
Ссылка: Чен, Дж. «Оценка индексов растительности и модифицированного простого отношения для бореальных приложений». Канадский журнал дистанционного зондирования 22 (1996): 229-242.
Нелинейный индекс (NLI)
Этот индекс предполагает, что взаимосвязь между многими индексами растительности и биофизическими параметрами поверхности является нелинейной. Он линеаризует отношения с параметрами поверхности, которые имеют тенденцию быть нелинейными.
Ссылка: Goel, N., and W. Qin. «Влияние архитектуры навеса на взаимосвязь между различными индексами растительности и LAI и Fpar: компьютерное моделирование». Remote Sensing Reviews 10 (1994): 309-347.
Нормализованный разностный индекс растительности (NDVI)
Этот индекс является мерой здоровой зеленой растительности. Сочетание формулировки с нормализованной разницей и использования областей с самым высоким поглощением и отражением хлорофилла делает его надежным в широком диапазоне условий. Однако он может насыщаться в условиях густой растительности, когда LAI становится высоким.
Значение этого индекса колеблется от -1 до 1. Обычный диапазон для зеленой растительности составляет от 0,2 до 0,8.
Ссылка: Роуз, Дж., Р. Хаас, Дж. Шелл и Д. Диринг. Мониторинг систем растительности на Великих равнинах с помощью ERTS . Третий симпозиум ERTS, НАСА (1973 г.): 309–317.
Оптимизированный индекс растительности с поправкой на почву (OSAVI)
Этот индекс основан на индексе растительности с поправкой на почву (SAVI). Он использует стандартное значение 0,16 для коэффициента корректировки фона купола. Рондо (1996) определили, что это значение обеспечивает большую изменчивость почвы, чем SAVI, для низкого растительного покрова, демонстрируя при этом повышенную чувствительность к растительному покрову более 50%. Этот индекс лучше всего использовать в районах с относительно редкой растительностью, где почва видна сквозь полог.
Ссылка: Рондо Г.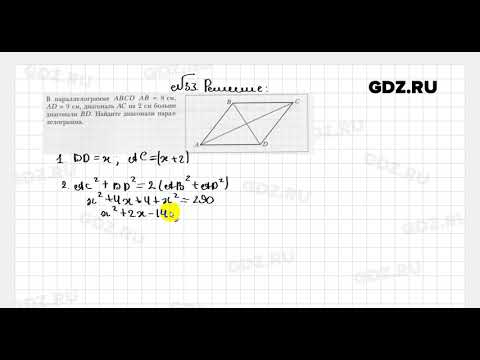 , М. Стивен и Ф. Барет. «Оптимизация индексов растительности с поправкой на почву». Дистанционное зондирование окружающей среды 55 (1996): 95-107.
, М. Стивен и Ф. Барет. «Оптимизация индексов растительности с поправкой на почву». Дистанционное зондирование окружающей среды 55 (1996): 95-107.
Перенормированный разностный индекс растительности (RDVI)
Этот индекс использует разницу между длинами волн в ближнем инфракрасном и красном диапазонах вместе с NDVI, чтобы выделить здоровую растительность. Он нечувствителен к влиянию почвы и геометрии обзора солнца.
Ссылка: Roujean, J., and F. Breon. «Оценка ФАР, поглощаемой растительностью, на основе измерений двунаправленного отражения». Дистанционное зондирование окружающей среды 51 (1995): 375-384.
Индекс растительности с поправкой на почву (SAVI)
Этот индекс аналогичен NDVI, но подавляет влияние пикселей почвы. Он использует поправочный коэффициент фона растительного покрова, L , который является функцией плотности растительности и часто требует предварительного знания количества растительности. Huete (1988) предлагает оптимальное значение 90 604 L 90 605 =0,5 для учета фоновых изменений почвы первого порядка. Этот индекс лучше всего использовать в районах с относительно редкой растительностью, где почва видна сквозь полог.
Huete (1988) предлагает оптимальное значение 90 604 L 90 605 =0,5 для учета фоновых изменений почвы первого порядка. Этот индекс лучше всего использовать в районах с относительно редкой растительностью, где почва видна сквозь полог.
Ссылка: Huete, A. «Индекс растительности с поправкой на почву (SAVI)». Дистанционное зондирование окружающей среды 25 (1988): 295-309.
Простой коэффициент (SR)
Этот индекс представляет собой отношение (1) длины волны с наивысшей отражательной способностью для растительности и (2) длины волны с самым глубоким поглощением хлорофилла. Простое уравнение легко понять, и оно эффективно в широком диапазоне условий. Как и в случае с NDVI, он может насыщаться густой растительностью, когда LAI становится очень высоким.
Ссылка: Рождение, Г. и Г. Маквей. «Измерение цвета растущего газона с помощью спектрофотометра отражения». Агрономический журнал 60 (1968): 640-643.
Суммарный индекс зелени (SGI)
Этот индекс является одним из простейших индексов растительности, используемых для обнаружения изменений в зелени растительности. Поскольку свет сильно поглощается зеленой растительностью в этой спектральной области, индекс SG очень чувствителен к небольшим изменениям раскрытия растительного полога, например, к тому, что может произойти при волнении леса.
SGI — это среднее значение коэффициента отражения в диапазоне от 500 до 600 нм спектра. Затем сумма нормализуется по количеству полос, чтобы преобразовать ее обратно в единицы отражательной способности.
Значение этого индекса находится в диапазоне от 0 до более чем 50 (в единицах % отражения). Обычный диапазон для зеленой растительности составляет от 10 до 25 процентов.
Ссылка: Лобелл Д. и Г. Аснер. «Hyperion Исследования стресса сельскохозяйственных культур в Мексике». Материалы 12-го ежегодного семинара JPL по бортовым наукам о Земле , Пасадена, Калифорния (2003 г.).
Материалы 12-го ежегодного семинара JPL по бортовым наукам о Земле , Пасадена, Калифорния (2003 г.).
Трансформированный разностный индекс растительности (TDVI)
Этот индекс полезен для мониторинга растительного покрова в городской среде. Он не насыщает, как NDVI и SAVI.
Ссылка: Баннари А., Х. Асалхи и П. Тейе. «Преобразованный индекс различия растительности (TDVI) для картирования растительного покрова» В материалах Proceedings of the Geoscience and Remote Sensing Symposium , IGARSS ’02, IEEE International, Volume 5 (2002).
Индекс треугольной зелени (TGI)
Этот индекс аппроксимирует площадь треугольника, ограничивающего спектр отражения листа, где вершины находятся в красной, зеленой и синей длинах волн.
Члены лямбда ( λ ) представляют центральные длины волн соответствующих полос. Термины Rho ( ρ ) представляют значения пикселей этих полос. Исходное уравнение TGI (Hunt et al, 2011) использовало 670 нм, 550 нм и 480 нм для центров красной, зеленой и синей длин волн с шириной полосы 10 нм. На следующем рисунке показан спектральный профиль ENVI для пикселя, представляющего здоровую зеленую растительность. На ней показан треугольник, ограничивающий спектр растительности.
Термины Rho ( ρ ) представляют значения пикселей этих полос. Исходное уравнение TGI (Hunt et al, 2011) использовало 670 нм, 550 нм и 480 нм для центров красной, зеленой и синей длин волн с шириной полосы 10 нм. На следующем рисунке показан спектральный профиль ENVI для пикселя, представляющего здоровую зеленую растительность. На ней показан треугольник, ограничивающий спектр растительности.
Авторы предлагают заменить узкие полосы широкими полосами и полосами цифровых камер. ENVI использует широкополосные определения для красной, зеленой и синей длин волн. В этом случае значения TGI могут быть ниже определений длины волны в узкой полосе частот. Это связано с тем, что усреднение сигнала в большей области длин волн приводит к уменьшению коэффициента отражения зеленого цвета и увеличению коэффициента отражения красного цвета.
TGI сильно коррелирует с содержанием хлорофилла в листьях. Значения TGI положительны, когда коэффициент отражения зеленого больше, чем линия между красной и синей вершинами. Это соответствует зеленой растительности. TGI является отрицательным, когда коэффициент отражения зеленого меньше, чем линия между красной и синей вершинами. Это соответствует таким особенностям, как красноземы.
Это соответствует зеленой растительности. TGI является отрицательным, когда коэффициент отражения зеленого меньше, чем линия между красной и синей вершинами. Это соответствует таким особенностям, как красноземы.
Поскольку этот индекс измеряет длины волн красного, зеленого и синего цветов, он подходит для использования с датчиками беспилотных летательных аппаратов (БПЛА) с RGB-камерами.
Ссылка: Хант, Э., К. Дотри, Дж. Эйтель и Д. Лонг. «Дистанционное определение содержания хлорофилла в листьях с использованием индекса видимой полосы». Агрономический журнал 103, № 4 (2011): 1090-1099.
Индекс устойчивости к видимой атмосфере (VARI)
Этот индекс основан на ARVI и используется для оценки доли растительности на сцене с низкой чувствительностью к атмосферным воздействиям.
Ссылка: Gitelson, A., et al. «Линии растительности и почвы в видимом спектральном пространстве: концепция и метод дистанционной оценки доли растительности». International Journal of Remote Sensing 23 (2002): 2537−2562. Этот индекс подобен NDVI, но он использует весовой коэффициент ( a ), чтобы уменьшить несоответствие между вкладами ближнего инфракрасного и красного сигналов в NDVI. высокая плотность растительности, когда NDVI превышает 0,6. NDVI имеет тенденцию выравниваться, когда доля растительности и индекс площади листьев (LAI) увеличиваются, тогда как WDRVI более чувствителен к более широкому диапазону фракций растительности и к изменениям LAI.
International Journal of Remote Sensing 23 (2002): 2537−2562. Этот индекс подобен NDVI, но он использует весовой коэффициент ( a ), чтобы уменьшить несоответствие между вкладами ближнего инфракрасного и красного сигналов в NDVI. высокая плотность растительности, когда NDVI превышает 0,6. NDVI имеет тенденцию выравниваться, когда доля растительности и индекс площади листьев (LAI) увеличиваются, тогда как WDRVI более чувствителен к более широкому диапазону фракций растительности и к изменениям LAI.
Весовой коэффициент ( a ) может принимать значения от 0,1 до 0,2. ENVI использует значение 0,2 в соответствии с рекомендациями Henebry, Viña и Gitelson (2004). Чтобы применить другое значение весового коэффициента, используйте инструмент Band Math.
Ссылки:
Гительсон, А. «Индекс растительности с широким динамическим диапазоном для удаленной количественной оценки биофизических характеристик растительности». Журнал физиологии растений 161, № 2 (2004): 165-173.
Журнал физиологии растений 161, № 2 (2004): 165-173.
Хенебри Г., А. Винья и А. Гительсон. «Индекс растительности с широким динамическим диапазоном и его потенциальная полезность для анализа пробелов». Gap Analysis Bulletin 12: 50-56.
Улучшенный вегетативный индекс WorldView (WV-VI)
Этот индекс использует полосы WorldView-2 для расчета NDVI.
Значение этого индекса колеблется от -1 до 1. Обычный диапазон для зеленой растительности составляет от 0,2 до 0,8.
Ссылка: Вольф, А. Использование изображений WorldView 2 Vis-NIR MSI для поддержки картографирования земель и извлечения признаков с использованием нормализованных соотношений индексов различий Неопубликованный отчет, Longmont, CO: DigitalGlobe (2010).
Связанные темы
Спектральные индексы, индексы растительности, инструменты анализа растительности, растительность и ее отражательные свойства, Учебное пособие по индексам растительности EO-1 Hyperion
Об оценке общей первичной продуктивности средиземноморских пастбищ при различных режимах удобрения с использованием индексов растительности и гиперспектральных коэффициент отражения
Айрес, Л. М., Пио, К. А., и Перейра, Дж. С.: Влияние засухи на энергию
и обмен водяного пара над Средиземным морем C 3 ∕ C 4 пастбища на юге Португалии, Agr. Лесной метеорол., 148, 565–579,
https://doi.org/10.1016/j.agrformet.2007.11.001, 2008a.
М., Пио, К. А., и Перейра, Дж. С.: Влияние засухи на энергию
и обмен водяного пара над Средиземным морем C 3 ∕ C 4 пастбища на юге Португалии, Agr. Лесной метеорол., 148, 565–579,
https://doi.org/10.1016/j.agrformet.2007.11.001, 2008a.
Айрес, Л. М., Пио, К. А., и Перейра, Дж. С.: Обмен углекислым газом над Средиземноморье C 3 ∕ C 4 пастбища в течение двух климатологически контрастные годы, Глоб. Изменить биол., 14, 539–555, https://doi.org/10.1111/j.1365-2486.2007.01507.x, 2008b.
Анскер, П., Дуру, М., Тео, Дж. П., и Круз, П.: Конвергенция растений черты между видами в пастбищных сообществах упрощает их мониторинг, экол. Indic., 9, 1020–1029, https://doi.org/10.1016/J.ECOLIND.2008.12.002, 2009.
Аснер, Г. П., Непстад, Д., Кардино, Г., и Рэй, Д.: Стресс от засухи и
поглощение углерода в лесу Амазонки, измеренное с помощью космических снимков
спектроскопия, П. Натл. акад. науч. США, 101, 6039–6044,
https://doi. org/10.1073/pnas.0400168101, 2004.
org/10.1073/pnas.0400168101, 2004.
Бальзароло, М., Весково, Л., Хаммерле, А., Джанель, Д., Папале, Д., Томеллери, Э., и Вольфарт, Г.: О взаимосвязи между гиперспектральная отражательная способность и обмен CO 2 в европейских горных луга, Биогеонауки, 12, 3089–3108, https://doi.org/10.5194/bg-12-3089-2015, 2015.
Барси, Дж., Ли, К., Кваран, Г., Маркхэм, Б., и Педельти, Дж.: The Spectral Отклик оперативного наземного тепловизора Landsat-8, дистанционное зондирование, 6, 10232–10251, https://doi.org/10.3390/rs61010232, 2014.
Бейтс, Д., Мэхлер, М., Болкер, Б., и Уокер, С.: Линейная подгонка Модели смешанных эффектов с использованием lme4, J. Stat. софт, 67, 1–48, https://doi.org/10.18637/jss.v067.i01, 2014.
Бир, К., Райхштейн, М., Томеллери, Э., Сиаис, П., Юнг, М., Карвалье, Н.,
Рёденбек, К., Араин, М. А., Бальдокки, Д., Бонан, Г. Б., Бондо, А.,
Ческатти А., Ласслоп Г., Линдрот А., Ломас М., Луиссарт С., Марголис,
Х., Олесон К.В., Рупсард О. , Венендал Э., Виови Н., Уильямс К.,
Вудворд, Ф. И., и Папейл, Д.: Валовое поглощение двуокиси углерода землей:
глобальное распространение и ковариация с климатом, Наука, 329, 834–838,
https://doi.org/10.1126/science.1184984, 2010.
, Венендал Э., Виови Н., Уильямс К.,
Вудворд, Ф. И., и Папейл, Д.: Валовое поглощение двуокиси углерода землей:
глобальное распространение и ковариация с климатом, Наука, 329, 834–838,
https://doi.org/10.1126/science.1184984, 2010.
Борер, Э. Т., Грейс, Дж. Б., Харпол, В. С., МакДугалл, А. С., и Сиблум, Э. У.: Десять лет изучения реакции пастбищных экосистем на глобальные изменение окружающей среды, Нац. Экол. Эволюция, 1, 0118, https://doi.org/10.1038/s41559-017-0118, 2017.
Бугало, М. Н. и Абреу, Дж. М.: Многофункциональная роль пастбищ, в: Устойчивые средиземноморские пастбища и их многофункциональность, Vol. 79, Option Méditerraneenes, под редакцией: Porqueddu, C. и Tavares de Соуза, М. М., CIHEAM/FAO/ENMP/SPPF, Сарагоса, 25–30, 2008 г.
Кадима Дж., Сердейра Дж. и Минхото М.: Вычислительные аспекты алгоритмов выбора переменных в контексте главных компонент, вычисл. Стат. Data An., 47, 225–236, https://doi.org/10.1016/j.csda.2003.11.001, 2004.
Casas, A. , Riaño, D., Ustin, S.L., Dennison, P. и Салас, Дж.:
Оценка связанных с водой биохимических и биофизических свойств растительности
использование мультивременных аэроспектральных данных и их сравнение с MODIS
спектральный отклик, Remote Sens. Environ., 148, 28–41,
https://doi.org/10.1016/j.rse.2014.03.011, 2014.
, Riaño, D., Ustin, S.L., Dennison, P. и Салас, Дж.:
Оценка связанных с водой биохимических и биофизических свойств растительности
использование мультивременных аэроспектральных данных и их сравнение с MODIS
спектральный отклик, Remote Sens. Environ., 148, 28–41,
https://doi.org/10.1016/j.rse.2014.03.011, 2014.
Черасоли С., Кампаньоло М., Фариа Дж., Ногейра С. и Калдейра М.С.: Гиперспектральная отражательная способность, газообмен и метеорологические условия на пастбищах участки с различными режимами внесения удобрений в Центральной Португалии с марта по июнь 2016 г., ПАНГЕЯ, https://doi.org/10.1594/PANGAEA.8, 2018.
Ceulemans, T., Stevens, C.J., Duchateau, L., Jacquemyn, H., Gowing, D.J.
Г., Меркс Р., Уоллес Х., ван Ройен Н., Гетхем Т., Боббинк Р.,
Дорланд Э., Гаудник К., Алар Д., Коркет Э., Мюллер С., Дизе Н. Б.,
Дюпре, К., Дикманн, М., и Хонней, О.: Ограничения содержания фосфора в почве
биоразнообразие европейских пастбищ, Glob. Change Biol., 20, 3814–3822,
https://doi. org/10.1111/gcb.12650, 2014 г.
org/10.1111/gcb.12650, 2014 г.
Ченг Ю.-Б., Чжан К., Ляпустин А.И., Ван Ю. и Миддлтон Э.М.: Влияние эффективности использования света и параметризации fPAR на общую первичную производственное моделирование, с/х. Лесной метеорол., 189–190, 187–197, https://doi.org/10.1016/j.agrformet.2014.01.006, 2014.
Коста, А. К., Сантос, Дж. А., и Пинто, Дж. Г.: Сценарии изменения климата для экстремальные осадки в Португалии, Теор. заявл. Климатол., 108, 217–234, https://doi.org/10.1007/s00704-011-0528-3, 2012.
Craine, J.M., Froehle, J., Tilman, D.G., Wedin, D.A., and Chapin, III, F. S.: Взаимосвязь корневых и листовых признаков 76 травянистых видов и растений. относительное обилие по градиентам фертильности и беспокойства, Oikos, 93, 274–285, https://doi.org/10.1034/j.1600-0706.2001.0.x, 2001.
Cristiano, P.M., Posse, G., Di Bella, C.M., and Jaimes, F.R.:
Неопределенности в оценке fPAR травяных покровов при различных нагрузках
ситуации и различия в архитектуре, Int. Дж. Дистанционный Сенсор, 31,
4095–4109, https://doi.org/10.1080/01431160
Дж. Дистанционный Сенсор, 31,
4095–4109, https://doi.org/10.1080/01431160
9192, 2010.
Дэш, Дж. и Карран, П.Дж.: Индекс земного хлорофилла MERIS, Int. Дж. Remote Sens., 25, 5403–5413, https://doi.org/10.1080/0143116042000274015, 2004.
Диас-Вилья, доктор медицинских наук, Мараньон, Т., Арройо, Дж., и Гарридо, Б.: Почвенный банк семян и флористическое разнообразие в мозаике лесов и лугов в юг Испании, J. Veg. наук, 14, 701–709, https://doi.org/10.1111/j.1654-1103.2003.tb02202.x, 2003.
Друш, М., Дель Белло, У., Карлье, С., Колин, О., Фернандес, В., Гаскон, Ф., Хёрш Б., Изола К., Лаберинти П., Мартимор П., Мейгрет А., Спото Ф., Сай, О., Марчезе, Ф. и Барджеллини, П.: Sentinel-2: Оптическая система ЕКА. Миссия с высоким разрешением для оперативных служб GMES, дистанционное зондирование окружающей среды, 120, 25–36, https://doi.org/10.1016/j.rse.2011.11.026, 2012.
ESA: Отчет о качестве данных Sentinel-2, выпуск 24 (февраль 2018 г.) — Sentinel-2
Библиотека документов MSI — Руководства пользователя — Sentinel Online, доступна по адресу:
https://earth. esa.int/web/sentinel/user-guides/sentinel-2-msi/document-library/-/asset_publisher/Wk0TKajiISaR/content/
(последний доступ: 25 января 2018 г.), 2018.
esa.int/web/sentinel/user-guides/sentinel-2-msi/document-library/-/asset_publisher/Wk0TKajiISaR/content/
(последний доступ: 25 января 2018 г.), 2018.
Феншолт, Р., Сандхолт, И., и Расмуссен, М.С.: Оценка MODIS LAI, fAPAR и взаимосвязь между fAPAR и NDVI в полузасушливой среде с использованием измерений на месте, Remote Sens. Environment, 91, 490–507, https://doi.org/10.1016/j.rse.2004.04.009, 2004.
Филелья, И., Пеньюэлас, Дж., Льоренс, Л., и Эстиарте, М.: Отражение оценка сезонных и годовых изменений биомассы и поглощения CO 2 средиземноморского кустарника, подвергшегося экспериментальному потеплению и засухе, Remote Sens. Environ., 90, 308–318, https://doi.org/10.1016/J.RSE.2004.01.010, 2004.
Филелла, И., Поркар-Кастелл, А., Мунне-Бош, С., Бэк, Дж.,
Гарбульский, М. Ф., и Пеньюэлас, Дж.: Оценка PRI долгосрочных изменений
соотношения каротиноиды/хлорофилл и кратковременные изменения деэпоксидации
состояние ксантофиллового цикла, Int. J. Remote Sens. , 30, 4443–4455,
https://doi.org/10.1080/01431160802575661, 2009 г..
, 30, 4443–4455,
https://doi.org/10.1080/01431160802575661, 2009 г..
Филиппа Г., Кремонезе Э., Мильявакка М., Гальваньо М., Форкель М., Вингейт Л., Томеллери Э., Морра ди Селла У. и Ричардсон А. Д.: Phenopix: пакет R для фенологии растительности на основе изображений, Agr. Лес Meteorol., 220, 141–150, https://doi.org/10.1016/j.agrformet.2016.01.006, 2016. Technometrics, 16, 499–511, https://doi.org/10.1080/00401706.1974.10489231, 1974. W., Seitzinger, S.P., Asner, G.P., Cleveland, C.C., Green, P.A., Holland, Э. А., Карл Д. М., Майклс А. Ф., Портер Дж. Х., Таунсенд А. Р. и Восмарти, CJ: Циклы азота: прошлое, настоящее и будущее, биогеохимия, 70, 153–226, https://doi.org/10.1007/s10533-004-0370-0, 2004.
Гамон, Дж. А.: Обзоры и синтезы: оптическая выборка магнитной башни след, Биогеонауки, 12, 4509–4523, https://doi.org/10.5194/bg-12-4509-2015, 2015.
Гамон, Дж. А., Пенуэлас, Дж., и Филд, С. Б.: Спектральный спектр в узком диапазоне волн.
индекс, который отслеживает суточные изменения эффективности фотосинтеза, Remote Sens. Environ., 41, 35–44, 1992.
Environ., 41, 35–44, 1992.
Гамон, Дж. А., Серрано, Л., и Серфус, Дж. С.: Фотохимическая отражательная способность индекс: оптический показатель эффективности использования фотосинтетического излучения через виды, функциональные типы и уровни питательных веществ, Oecologia, 112, 492–501, https://doi.org/10.1007/s004420050337, 1997.
Гао, Б.: NDWI – Нормализованный разностный индекс воды для дистанционного зондирования растительность жидкая вода из космоса, Remote Sens. Environ., 58, 257–266, https://doi.org/10.1016/S0034-4257(96)00067-3, 1996.
Гарбульский М.Ф., Пеньюэлас Дж., Гамон Дж., Иноуэ Ю. и Филелла И.: Фотохимический индекс отражения (PRI) и дистанционное зондирование листа, Эффективность использования излучения растительного покрова и экосистем Обзор и метаанализ, Удаленный датчик окружающей среды, 115, 281–29.7, https://doi.org/10.1016/j.rse.2010.08.023, 2011.
Джанель Д., Весково Л., Марколла Б., Манка Г. и Ческатти А.:
Потоки углерода в экосистеме и спектральная отражательная способность полога горного луга,
Междунар. J. Remote Sens., 30, 435–449, https://doi.org/10.1080/01431160802314855, 2009.
J. Remote Sens., 30, 435–449, https://doi.org/10.1080/01431160802314855, 2009.
Гительсон, А. и Мерзляк, М. Н.: Изменения спектрального отражения, связанные с с осенним старением Aesculus-Hippocastanum L. и Листья Acer-Platanoides L – спектральные характеристики и связь с оценка хлорофилла, J. Plant Physiol., 143, 286–29.2, 1994.
Гительсон А. А. и Мерзляк М. Н.: Дистанционное зондирование хлорофилла концентрация в листьях высших растений, Adv. Космические исследования, 22, 689–692, https://doi.org/10.1016/S0273-1177(97)01133-2, 1998.
Гительсон А.А., Кауфман Ю.Дж. и Мерзляк М.Н.: Использование зеленого канала в дистанционном зондировании глобальной растительности с помощью EOS-MODIS, Remote Sens. Environ., 58, 289–298, https://doi.org/10.1016/S0034-4257(96)00072-7, 1996.
Гительсон, А. А., Гамон, Дж. А., и Соловченко, А.: Множественные драйверы
сезонные изменения в PRI: последствия для фотосинтеза 1. Уровень листа, удаленный
Сан. Экология, 191, 110–116, https://doi.org/10.1016/j.rse.2016.12.014, 2017а.
Экология, 191, 110–116, https://doi.org/10.1016/j.rse.2016.12.014, 2017а.
Гительсон, А. А., Гамон, Дж. А., и Соловченко, А.: Несколько водителей сезонные изменения PRI: последствия для фотосинтеза 2. Уровень насаждения, Remote Sens. Environ., 190, 198–206, https://doi.org/10.1016/J.RSE.2016.12.015, 2017b.
Гленн Э., Хьюте А., Наглер П. и Нельсон С.: отношения между Индексы растительности с помощью дистанционного зондирования, атрибуты растительного покрова и физиология растений Процессы: что индексы растительности могут и не могут рассказать нам о Пейзаж, Датчики, 8, 2136–2160, https://doi.org/10.3390/s8042136, 2008.
Гауэр С.Т., Кучарик С.Дж. и Норман Дж.М.: Прямое и косвенное Оценка индекса листовой поверхности, fAPAR и чистой первичной продукции Наземные экосистемы, дистанционное зондирование окружающей среды, 70, 29–51, https://doi.org/10.1016/S0034-4257(99)00056-5, 1999.
Харпол, В.С., Поттс, Д.Л., и Судинг, К.Н.: Реакция экосистемы на воду
и азотная поправка на лугах Калифорнии, Glob. Смена биол., 13,
2341–2348, https://doi.org/10.1111/j.1365-2486.2007.01447.x, 2007.
Смена биол., 13,
2341–2348, https://doi.org/10.1111/j.1365-2486.2007.01447.x, 2007.
Хотхорн Т., Бретц Ф. и Вестфолл П.: Общий вывод об одновременном выводе Параметрические модели, Biometrical J., 50, 346–363, https://doi.org/10.1002/bimj.200810425, 2008.
INMG: О климате Португалии. Normais climatológicas da região Ribatejo e Oeste корреспонденты a 1951–1980, Instituto Nacional de Meteorologia e Geofisica, Лиссабон, Португалия, 1991. Дж., Банза Дж., Дэвид Дж., Мильетта Ф., Боргетти М., Манка Г. и Валентини, Р.: Высыхание и увлажнение средиземноморских почв стимулирует разложение и эмиссия углекислого газа: «эффект березы», Физиология дерева, 27, 929–940, https://doi.org/10.1093/treephys/27.7.929, 2007.
Джоэл Г., Гамон Дж. А. и Филд С. Б.: Эффективность производства подсолнечника: Роль водного и азотного стресса, Remote Sens. Environ., 62, 176–188, https://doi.org/10.1016/s0034-4257(97)00093-x, 1997.
Джонс, Х.Г.: Растения и микроклимат: количественный подход к
Экологическая физиология растений, 2-е изд. , издательство Кембриджского университета,
Cambridge, 1992.
, издательство Кембриджского университета,
Cambridge, 1992.
Jongen, M., Pereira, J.S., Aires, L.M., и Pio, C.A.: Эффекты засухи и времени выпадения осадков на межгодовой ход обмен экосистема-атмосфера на средиземноморских пастбищах, Agr. Лес Метеорол., 151, 595–606, https://doi.org/10.1016/j.agrformet.2011.01.008, 2011.
Jongen, M., Lecomte, X., Unger, S., Fangueiro, D. и Pereira, J.S. : Изменчивость осадков не влияет на дыхание почвы и азот. динамика подлеска средиземноморского дуба, Plant Soil, 372, 235–251, https://doi.org/10.1007/s11104-013-1728-7, 2013a.
Йонген М., Унгер С., Фангейро Д., Черасоли С., Сильва Х.М.Н. и Перейра, Дж. С.: Устойчивость нижнего яруса монтадо к экспериментальным изменчивость осадков нарушается в условиях сильной естественной засухи, Agr. Экосистем. Окружающая среда., 178, 18–30, 2013б.
Кира, О., Нгуи-Робертсон, А.Л., Аркебауэр, Т.Дж., Линкер, Р., и Гительсон,
А. А.: На пути к универсальным моделям для оценки зеленого LAI для кукурузы и сои:
Спутниковые наблюдения, Remote Sens. , 9, 318, https://doi.org/10.3390/rs
, 9, 318, https://doi.org/10.3390/rs
Корхонен, Л., Хади, Пакален, П., и Раутиайнен, М.: Сравнение Sentinel-2 и Landsat-8 в оценке полога бореальных лесов и индекс площади листа, Remote Sens. Environ., 195, 259–274, https://doi.org/10.1016/j.rse.2017.03.021, 2017.
Ковац Р.С., Валентини Р., Бауэр Л.М., Георгопулу Э., Якоб Д., Мартин Э., Раунсевелл М. и Суссана Дж. Ф.: Европа, в: Изменение климата. 2014: Воздействие, адаптация и уязвимость. Часть B: Региональные аспекты. Вклад Рабочей группы II в пятый доклад об оценке Межправительственная группа экспертов по изменению климата, 1267–1326 гг., Кембриджский университет. Press, Cambridge, UK and New York, NY, USA, 2014.
Лавленд, Т. Р. и Айронс, Дж. Р.: Landsat-8: Планы, наследие, Remote Sens. Environment, 185, 1–6, https://doi.org/10.1016/j.rse.2016.07.033, 2016.
Ламли, Т .: Томас Ламли с использованием кода Фортрана Алана Миллера (2009). Прыжки:
выбор подмножества регрессии, версия пакета R 2.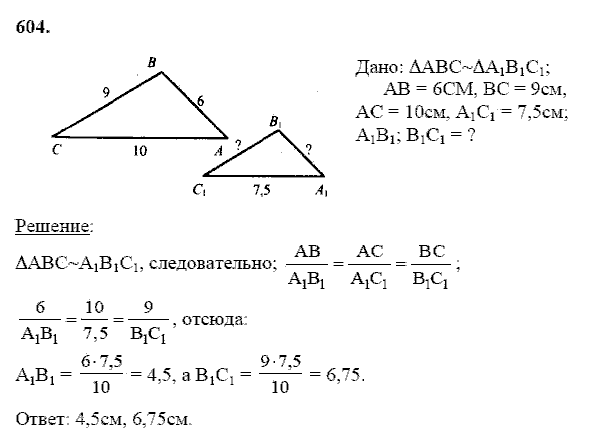 9, 2009 г.
и Бальдокки, Д.: Прогнозирование потока CO 2 в ландшафтном масштабе на пастбище.
и рисовые поля с долгосрочными гиперспектральными измерениями отражательной способности растительного покрова,
Биогеонауки, 12, 4577–4594, https://doi.org/10.5194/bg-12-4577-2015,
2015.
9, 2009 г.
и Бальдокки, Д.: Прогнозирование потока CO 2 в ландшафтном масштабе на пастбище.
и рисовые поля с долгосрочными гиперспектральными измерениями отражательной способности растительного покрова,
Биогеонауки, 12, 4577–4594, https://doi.org/10.5194/bg-12-4577-2015,
2015.
Мерзляк М.Н., Гительсон А.А., Чивкунова О.Б., Ракитин В.Ю.: Неразрушающее оптическое обнаружение изменений пигмента во время старения листьев и созревание плодов, Physiol. Завод., 106, 135–141, https://doi.org/10.1034/j.1399-3054.1999.106119.x, 1999.
Monteith, J.L.: Solar Radiation and Productivity in Tropical Ecosystems, J. заявл. Ecol., 9, 747–766, https://doi.org/10.2307/2401901, 1972.
Монтейт, Дж. Л.: Климат и эффективность растениеводства в Великобритании, Филос. Т. Рой. соц. B, 281, 277–294, 1977.
Морено, А., Маселли, Ф., Жилаберт, М.А., Кьези, М., Мартинес, Б., и
Зойферт, Г.: Оценка изображений MODIS для отслеживания эффективности использования света в
маловодный средиземноморский сосновый бор, Remote Sens. Environ., 123,
359–367, https://doi.org/10.1016/j.rse.2012.04.003, 2012.
Environ., 123,
359–367, https://doi.org/10.1016/j.rse.2012.04.003, 2012.
Минени, Р. Б. и Уильямс, Д. Л.: Об отношениях между FAPAR и NDVI, Remote Sens. Environ., 49, 200–211, 1994.
Нестола, Э., Кальфапьетра, К., Эммертон, К. и Гамон, Дж. А.: Мониторинг сезонной динамики углерода на пастбищах, интегрируя MODIS NDVI, проксимальную оптическую выборку и вихревую ковариацию измерения, Remote Sens., 8, 260, https://doi.org/10.3390/rs8030260, 2016.
Отани, К.: Начальная загрузка R 2 и скорректированный R 2 в регрессии анализ, экон. Модель., 17, 473–483, https://doi.org/10.1016/S0264-9993(99)00034-6, 2000.
Оллингер, С.В.: Источники изменчивости коэффициента отражения растительного покрова и конвергентные свойства растений, New Phytol., 189, 375–394, https://doi.org/10.1111/j.1469-8137.2010.03536.x, 2011.
Пеньюэлас, Дж., Филелла, И., Биль, К., Серрано, Л., и Саве, Р.:
отражение на 9область 50–970 нм как индикатор состояния воды растений,
Междунар.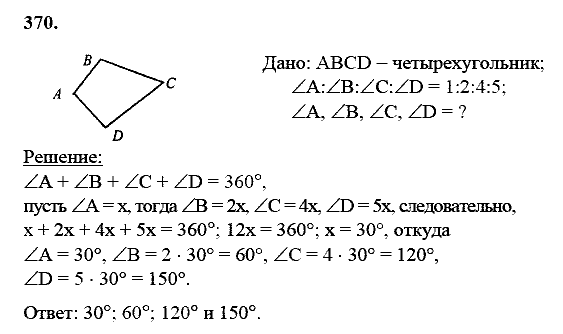 J. Remote Sens., 14, 1887–1905, https://doi.org/10.1080/0143116
J. Remote Sens., 14, 1887–1905, https://doi.org/10.1080/0143116
54010, 1993. эффективность использования излучения со спектральной отражательной способностью, New Phytol., 131, 291–296, https://doi.org/10.1111/j.1469-8137.1995.tb03064.x, 1995.
Пеньюэлас, Дж., Пинол, Дж., Огая, Р., и Филелла, И.: Оценка растения концентрация воды по коэффициенту отражения Water Index WI (R900/R970), внутр. Дж. Remote Sens., 18, 2869–2875, https://doi.org/10.1080/014311697217396, 1997.
Peñuelas, J., Poulter, B., Sardans, J., Ciais, P., van der Velde, М., Бопп Л., Буше О., Годдерис Ю., Хинсингер П., Льюсия Дж., Нардин Э., Викка, С., Оберштайнер, М., и Янссенс, И. А.: Индуцированные человеком дисбаланс азота и фосфора изменяет естественные и управляемые экосистемы по всему миру. глобус, нац. Commun., 4, 2934, https://doi.org/10.1038/ncomms3934, 2013.
Перес-Приего, О., Гуан, Дж., Россини, М., Фава, Ф., Вутцлер, Т., Морено,
Г., Карвалье Н., Каррара А., Колле О., Юлитта Т., Шрампф М. ,
Рейхштейн, М., и Мильявакка, М.: Индуцированная солнцем флуоресценция хлорофилла и
индекс фотохимической отражательной способности улучшает дистанционное зондирование
оценки производства при различной доступности питательных веществ в типичном
Экосистема средиземноморской саванны, Biogeosciences, 12, 6351–6367,
https://doi.org/10.5194/бг-12-6351-2015, 2015а.
,
Рейхштейн, М., и Мильявакка, М.: Индуцированная солнцем флуоресценция хлорофилла и
индекс фотохимической отражательной способности улучшает дистанционное зондирование
оценки производства при различной доступности питательных веществ в типичном
Экосистема средиземноморской саванны, Biogeosciences, 12, 6351–6367,
https://doi.org/10.5194/бг-12-6351-2015, 2015а.
Перес-Приего, О., Лопес-Бальестерос, А., Санчес-Каньете, Э., Серрано-Ортис П., Куцбах Л., Доминго Ф., Югстер В. и Ковальски А.: Анализ неопределенностей при расчете потоков с использованием камеры: случайные и систематические ошибки, Plant Soil, 393, 229–244, https://doi.org/10.1007/s11104-015-2481-x, 2015b.
Поркар-Кастелл, А., Гарсия-Плазаола, Дж. И., Николь, С. Дж., Колари, П.,
Оласкоага Б., Куусинен Н., Фернандес-Марин Б., Пулккинен М., Юрола Э.,
и Никинмаа, Э.: Физиология сезонных отношений между
индекс фотохимического отражения и эффективность использования фотосинтетического света,
Экология, 170, 313–323, https://doi. org/10.1007/s00442-012-2317-9, 2012.
org/10.1007/s00442-012-2317-9, 2012.
Поркар-Кастелл, А., Мак Артур, А., Россини, М., Эклунд, Л., Пачеко-Лабрадор Дж., Андерсон К., Бальсароло М., Мартин М.П., Джин, Х., Томеллери Э., Черазоли С., Саковска К., Хуэни А., Юлитта Т., Никол, С. Дж., и Весково, Л.: EUROSPEC: на стыке между дистанционное зондирование и экосистема CO 2 измерения потоков в Европе, Биогеонауки, 12, 6103–6124, https://doi.org/10.5194/bg-12-6103-2015, 2015.
R Основная группа: R: язык и среда для статистических вычислений, R Foundation for Statistical Computing, Вена, Австрия, доступно по адресу: https://www.R-project.org/ (последний доступ: 7 августа 2018 г.), 2017 г.
Райхштейн М., Рей А., Фрейбауэр А., Тенхунен Дж., Валентини Р., Банза,
Дж., Казальс П., Ченг Ю., Грюнцвейг Дж. М., Ирвин Дж., Жоффр Р., Лоу,
Б. Э., Лустау Д., Мильетта Ф., Эхель В., Урсиваль Ж.-М., Перейра Ж.
С., Перессотти А., Понти Ф., Ци Ю., Рамбал С., Раймент М., Романя Дж.,
Росси Ф., Тедески В., Тироне Г. , Сюй М. и Якир Д.: Моделирование временных
и крупномасштабная пространственная изменчивость дыхания почвы из почвенной воды
индексы доступности, температуры и продуктивности растительности, глобальные
Биогеохим. с., 17, 1104, https://doi.org/10.1029/2003GB002035, 2003.
, Сюй М. и Якир Д.: Моделирование временных
и крупномасштабная пространственная изменчивость дыхания почвы из почвенной воды
индексы доступности, температуры и продуктивности растительности, глобальные
Биогеохим. с., 17, 1104, https://doi.org/10.1029/2003GB002035, 2003.
Россини, М., Коглиати, С., Мерони, М., Мильявакка, М., Гальваньо, М., Бусетто, Л., Кремонезе, Э., Хулитта, Т., Синискалко, К., Морра ди Селла, У., и Коломбо, Р.: Оценка валовой первичной продукции на основе дистанционного зондирования. на субальпийских пастбищах, Biogeosciences, 9, 2565–2584, https://doi.org/10.5194/bg-9-2565-2012, 2012.
Роуз, Дж. В., Хаас, Р. Х., Шелл, Дж. А., и Диринг, Д. У.: Мониторинг системы растительности на великих равнинах с ERTS, в: Третий симпозиум ERTS, Том. 1, СП-351, 309–317, НАСА, Вашингтон, округ Колумбия, США, 1974.
Саковска К., Весково Л., Марколла Б., Ющак Р., Олейник Дж. и
Джанель, Д.: Мониторинг потоков углекислого газа на субальпийских пастбищах.
экосистемы итальянских Альп с помощью мультиспектрального датчика Biogeosciences,
11, 4695–4712, https://doi. org/10.5194/bg-11-4695-2014, 2014.
org/10.5194/bg-11-4695-2014, 2014.
Саковска К., Ющак Р. и Джанель Д.: Дистанционное зондирование пастбищ Биофизические параметры в контексте спутниковой миссии Sentinel-2, Дж. Датчики, 2016, 1–16, https://doi.org/10.1155/2016/4612809., 2016.
Сала, О. Э.: Сценарии глобального биоразнообразия на 2100 год, Наука, 287, 1770–1774 гг., https://doi.org/10.1126/science.287.5459.1770. 2000.
Шенк, Х. Дж. и Джексон, Р. Б.: Глубина укоренения, распространение боковых корней и подземные/надземные аллометрии растений в водоограниченных экосистемах, J. Ecol., 90, 480–494, https://doi.org/10.1046/j.1365-2745.2002.00682.x, 2002.
Шимель, Д., Павлик, Р., Фишер, Дж. Б., Аснер , Г. П., Саатчи С., Таунсенд, П., Миллер К., Франкенберг К., Хиббард К. и Кокс П.: Наблюдение наземные экосистемы и круговорот углерода из космоса, Глоб. Изменить биол., 21, 1762–1776, https://doi.org/10.1111/gcb.12822, 2015.
Сиблум Э. У., Борер Э. Т., Бакли Ю., Клеланд Э. Э., Дэвис К. , Фирн,
Дж., Харпол, В.С., Отье, Ю., Линд, Э., МакДугалл, А., Оррок, Дж.Л.,
Пробер, С. М., Адлер, П., Альберти, Дж., Майкл Андерсон, Т., Баккер, Дж. Д.,
Бидерман Л. А., Блюменталь Д., Браун К. С., Брудвиг Л. А., Калдейра М.,
Чу, К., Кроули, М.Дж., Далео, П., Дамшен, Э.И., Д’Антонио, К.М.,
Декраппео, Н. М., Дикман, Ч. Р., Ду, Г., Фэй, П. А., Фратер, П., Грюнер, Д.
С., Хагена Н., Гектор А., Хелм А., Хиллебранд Х., Хофмокель К. С.,
Хамфрис, Х.К., Ирибарн, О., Джин, В.Л., Кей, А., Киркман, К.П., Клейн,
Дж. А., Нопс, Дж. М. Х., Ла Пьер, К. Дж., Ладвиг, Л. М., Ламбринос, Дж. Г.,
Лики, А.Д.Б., Ли, К., Ли, В., Маккалли, Р., Мельбурн, Б., Митчелл, К.
Э., Мур, Дж. Л., Морган, Дж., Мортенсен, Б., О’Халлоран, Л. Р., Партель,
М., Паскуаль, Дж., Пайк, Д.А., Риш, А.С., Салгуэро-Гомес, Р., Санкаран,
М., Шютц М., Симонсен А., Смит М., Стивенс К., Салливан Л., Уордл,
Г. М., Волкович Э. М., Рагг П. Д., Райт Дж. и Ян Л.: Прогнозирование
Вторжение в пастбищные экосистемы: экзотическое доминирование — настоящее затруднение
богатства?, Глоб.
, Фирн,
Дж., Харпол, В.С., Отье, Ю., Линд, Э., МакДугалл, А., Оррок, Дж.Л.,
Пробер, С. М., Адлер, П., Альберти, Дж., Майкл Андерсон, Т., Баккер, Дж. Д.,
Бидерман Л. А., Блюменталь Д., Браун К. С., Брудвиг Л. А., Калдейра М.,
Чу, К., Кроули, М.Дж., Далео, П., Дамшен, Э.И., Д’Антонио, К.М.,
Декраппео, Н. М., Дикман, Ч. Р., Ду, Г., Фэй, П. А., Фратер, П., Грюнер, Д.
С., Хагена Н., Гектор А., Хелм А., Хиллебранд Х., Хофмокель К. С.,
Хамфрис, Х.К., Ирибарн, О., Джин, В.Л., Кей, А., Киркман, К.П., Клейн,
Дж. А., Нопс, Дж. М. Х., Ла Пьер, К. Дж., Ладвиг, Л. М., Ламбринос, Дж. Г.,
Лики, А.Д.Б., Ли, К., Ли, В., Маккалли, Р., Мельбурн, Б., Митчелл, К.
Э., Мур, Дж. Л., Морган, Дж., Мортенсен, Б., О’Халлоран, Л. Р., Партель,
М., Паскуаль, Дж., Пайк, Д.А., Риш, А.С., Салгуэро-Гомес, Р., Санкаран,
М., Шютц М., Симонсен А., Смит М., Стивенс К., Салливан Л., Уордл,
Г. М., Волкович Э. М., Рагг П. Д., Райт Дж. и Ян Л.: Прогнозирование
Вторжение в пастбищные экосистемы: экзотическое доминирование — настоящее затруднение
богатства?, Глоб.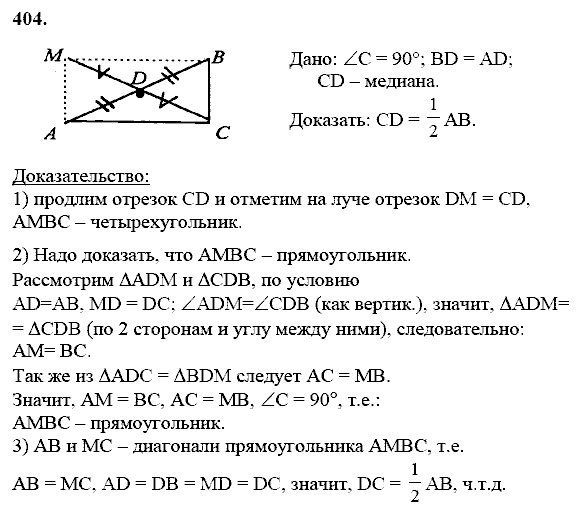 Изменить биол., 19, 3677–3687, https://doi.org/10.1111/gcb.12370,
2013.
Изменить биол., 19, 3677–3687, https://doi.org/10.1111/gcb.12370,
2013.
Tagesson, T., Fensholt, R., Huber, S., Horion, S., Guiro, I., Ehammer, A., и Ардо, Дж.: Определение сезонной динамики свойств экосистемы полузасушливые пастбища саванны из гиперспектральной отражательной способности на месте, Биогеонауки, 12, 4621–4635, https://doi.org/10.5194/bg-12-4621-2015, 2015.
Томас, Х.: Старение, старение и гибель всего растения, New Phytol., 197, 696–711, https://doi.org/10.1111/nph.12047, 2013.
Весково Л., Вольфарт Г., Бальцароло М., Пиллони С., Соттокорнола М., Родегьеро, М., и Джанель, Д.: Новые спектральные индексы растительности на основе плечевые длины волн ближнего инфракрасного диапазона для дистанционного обнаружения пастбищ фитомасса, межд. J. Remote Sens., 33, 2178–2195, https://doi.org/10.1080/01431161.2011.607195, 2012.
Викка С., Бальцароло М., Филелла И., Гранье А., Хербст М., Кноль А.,
Лонгдоз Б., Мунд М., Надь З., Пинтер К., Рамбал С., Вербесселт Дж. ,
Вергер, А., Зейлейс, А., Чжан, К., и Пеньюэлас, Дж.: Дистанционное зондирование
выявление влияния экстремальных засух на валовую первичную продукцию // Науч.
Респ.-УК, 6, 28269, https://doi.org/10.1038/srep28269, 2016.
,
Вергер, А., Зейлейс, А., Чжан, К., и Пеньюэлас, Дж.: Дистанционное зондирование
выявление влияния экстремальных засух на валовую первичную продукцию // Науч.
Респ.-УК, 6, 28269, https://doi.org/10.1038/srep28269, 2016.
Винья, А. и Гительсон, А.А.: Новые разработки в дистанционной оценке доли поглощенной фотосинтетически активной радиации сельскохозяйственными культурами, Геофиз. Рез. Lett., 32, L17403, https://doi.org/10.1029/2005GL023647, 2005.
Weisser, W.W., Roscher, C., Meyer, S.T., Ebeling, A., Luo, G., Allan, E. ,
Бесслер, Х., Барнард, Р.Л., Бухманн, Н., Бускот, Ф., Энгельс, К.,
Фишер, К., Фишер, М., Гесслер, А., Глейкснер, Г., Галле, С., Хильдебрандт,
А., Хиллебранд, Х., де Крон, Х., Ланге, М., Леймер, С., Ле Ру, X., Милку,
А., Моммер Л., Никлаус П.А., Ольманн Ю., Пру Р., Рой Дж., Шербер,
К., Шерер-Лоренцен М., Шой С., Чарнтке Т., Вахендорф М., Вагг,
К., Вайгельт А., Вилке В., Вирт К., Шульце Э.-Д., Шмид Б. и
Эйзенхауэр, Н.: Влияние биоразнообразия на функционирование экосистемы за 15 лет. эксперимент на пастбищах: закономерности, механизмы и открытые вопросы, Basic Appl.
Ecol., 23, 1–73, https://doi.org/10.1016/j.baae.2017.06.002, 2017.
эксперимент на пастбищах: закономерности, механизмы и открытые вопросы, Basic Appl.
Ecol., 23, 1–73, https://doi.org/10.1016/j.baae.2017.06.002, 2017.
Вестоби, М., Фальстер, Д. С., Молес, А. Т., Веск, П. А., и Райт, И. Дж.: Экологические стратегии растений: некоторые основные аспекты различий между Виды, Анну. Преподобный Экол. систем, 33, 125–159, https://doi.org/10.1146/annurev.ecolsys.33.010802.150452, 2002.
WRB: Рабочая группа IUSS: Всемирная справочная база почвенных ресурсов, 2006 г., 2-е место. Изд., Доклады о мировых почвенных ресурсах № 103, ФАО, Рим, Италия, 2006 г. над средиземноморским однолетним пастбищем в Калифорнии, Agr. Лесной метеорол., 123, 79–96, https://doi.org/10.1016/j.agrformet.2003.10.004, 2004.
Юань, В., Цай, В., Ся, Дж., Чен, Дж., Лю, С., Донг В., Мерболд Л., Лоу,
Б., Арайн А., Берингер Дж., Бернхофер К., Блэк А., Бланкен П. Д.,
Ческатти А., Чен Ю., Франсуа Л., Джанель Д., Янссенс И. А., Юнг,
М., Като Т., Кили Г., Лю Д., Марколла Б. , Монтаньяни Л., Раски А.,
Рупсар О., Варлагин А. и Вольфарт Г.: Глобальное сравнение использования света
модели эффективности для имитации валовой первичной обработки наземной растительности
производство на основе базы данных LaThuile, Agr. Лесной метеорол., 192–193,
108–120, https://doi.org/10.1016/j.agrformet.2014.03.007, 2014.
, Монтаньяни Л., Раски А.,
Рупсар О., Варлагин А. и Вольфарт Г.: Глобальное сравнение использования света
модели эффективности для имитации валовой первичной обработки наземной растительности
производство на основе базы данных LaThuile, Agr. Лесной метеорол., 192–193,
108–120, https://doi.org/10.1016/j.agrformet.2014.03.007, 2014.
Чжан, Ф. и Чжоу, Г.: Разработка алгоритма оценки эффективности использования света с использованием измерений гиперспектральной и вихревой ковариации in situ для кукурузы навес в Северо-Восточном Китае, Ecol. Эволюция, 7, 4735–4744, https://doi.org/10.1002/ece3.3051, 2017.
Межмасштабное взаимодействие размера дерева-хозяина и климатического дефицита воды определяет гибель деревьев, вызванную короедом
Резюме
Недавняя жаркая засуха в Калифорнии (2012–2016 гг.) привела к беспрецедентному выпадению сосны пондероза ( Pinus ponderosa ) смертность, в основном связанная с западным сосновым жуком ( Dendroctonus brevicomis ; WPB). Широкомасштабные климатические условия могут напрямую формировать модели смертности деревьев, но показатели смертности нелинейно реагируют на климат, когда характеристики леса в локальном масштабе влияют на поведение жуков-короедов (например, WPB). Чтобы проверить эти межмасштабные взаимодействия, мы проводим аэрофотосъемку на 32 участках вдоль градиента климатического дефицита воды (CWD), охватывающего 350 км по широте и 1000 м над уровнем моря в лесах Сьерра-Невады, затронутых WPB. Мы картируем, измеряем и классифицируем более 450 000 деревьев в пределах 9км 2 , проверка измерений с совпадающими полевыми графиками. Мы обнаружили, что больший размер, пропорция и плотность сосны пондероза (хозяин WPB) увеличивают уровень смертности хозяев, как и больший CWD. Важно отметить, что мы находим такое взаимодействие CWD/размера хозяина, что более крупные деревья увеличивают уровень смертности хозяев в жарких/сухих местах. Стратегии управления адаптацией к изменению климата должны учитывать, как беспокойство короедов может зависеть от межмасштабных взаимодействий, которые бросают вызов нашей способности прогнозировать и понимать закономерности гибели деревьев.
Широкомасштабные климатические условия могут напрямую формировать модели смертности деревьев, но показатели смертности нелинейно реагируют на климат, когда характеристики леса в локальном масштабе влияют на поведение жуков-короедов (например, WPB). Чтобы проверить эти межмасштабные взаимодействия, мы проводим аэрофотосъемку на 32 участках вдоль градиента климатического дефицита воды (CWD), охватывающего 350 км по широте и 1000 м над уровнем моря в лесах Сьерра-Невады, затронутых WPB. Мы картируем, измеряем и классифицируем более 450 000 деревьев в пределах 9км 2 , проверка измерений с совпадающими полевыми графиками. Мы обнаружили, что больший размер, пропорция и плотность сосны пондероза (хозяин WPB) увеличивают уровень смертности хозяев, как и больший CWD. Важно отметить, что мы находим такое взаимодействие CWD/размера хозяина, что более крупные деревья увеличивают уровень смертности хозяев в жарких/сухих местах. Стратегии управления адаптацией к изменению климата должны учитывать, как беспокойство короедов может зависеть от межмасштабных взаимодействий, которые бросают вызов нашей способности прогнозировать и понимать закономерности гибели деревьев.
Введение
Короеды нанесли последний удар многим из почти 150 миллионов деревьев, погибших во время жаркой засухи в Калифорнии 2012–2016 годов и ее последствий 1 . Предвестник грядущих последствий изменения климата, рекордно высокие температуры усугубили засуху 2,3 , которая усилила водный стресс у деревьев 4,5 , сделав их более восприимчивыми к заселению короедом 6,7 . Кроме того, столетие подавления пожаров позволило лесам превратиться в густые насаждения, что также может сделать их более уязвимыми для жуков-короедов 6,8,9 . Такое сочетание условий окружающей среды и структурных характеристик леса привело к гибели деревьев беспрецедентного масштаба по всему штату 10,11 .
Отмирание деревьев демонстрировало сильный градиент широты и высоты 4,11 , который можно лишь частично объяснить грубыми измерениями условий окружающей среды (т. е. исторического климатического дефицита воды; CWD) и современной структурой леса (т. площадь) 11 . Прогрессирующая потеря содержания воды в древесном покрове дает дополнительное представление о стрессе и риске гибели деревьев, но не может в конечном итоге определить, какие деревья на самом деле погибают от жуков-короедов, или выяснить факторы, влияющие на динамику популяции жуков-короедов и их распространение 5 . Короеды реагируют на локальные характеристики леса положительными обратными связями, которые нелинейно изменяют динамику гибели деревьев на фоне стрессовых условий окружающей среды 12,13 . Таким образом, явный учет местной структуры и состава леса 14,15 , а также его кросс-масштабное взаимодействие с региональными климатическими условиями 16 могут уточнить наше понимание закономерностей гибели деревьев в результате недавней жаркой засухи в Калифорнии. Задача одновременного измерения воздействия как локальных особенностей леса (таких как структура и состав), так и широкомасштабных условий окружающей среды (например, CWD) на нарушение леса насекомыми оставляет их эффект взаимодействия относительно малоизученным 14,15,16,17 .
площадь) 11 . Прогрессирующая потеря содержания воды в древесном покрове дает дополнительное представление о стрессе и риске гибели деревьев, но не может в конечном итоге определить, какие деревья на самом деле погибают от жуков-короедов, или выяснить факторы, влияющие на динамику популяции жуков-короедов и их распространение 5 . Короеды реагируют на локальные характеристики леса положительными обратными связями, которые нелинейно изменяют динамику гибели деревьев на фоне стрессовых условий окружающей среды 12,13 . Таким образом, явный учет местной структуры и состава леса 14,15 , а также его кросс-масштабное взаимодействие с региональными климатическими условиями 16 могут уточнить наше понимание закономерностей гибели деревьев в результате недавней жаркой засухи в Калифорнии. Задача одновременного измерения воздействия как локальных особенностей леса (таких как структура и состав), так и широкомасштабных условий окружающей среды (например, CWD) на нарушение леса насекомыми оставляет их эффект взаимодействия относительно малоизученным 14,15,16,17 .
Сосновые/смешанные хвойные леса пондероза в калифорнийском регионе Сьерра-Невада характеризуются регулярными нарушениями короедов, в первую очередь влиянием западного соснового лубоеда ( Dendroctonus brevicomis ; WPB) на сосну-хозяин пондероза ( Pinus ponderosa ) 18 . WPB — это первичный жук-короед: его репродуктивный успех зависит от смертности дерева-хозяина, что само по себе требует достаточного количества жуков для массовой атаки дерева-хозяина и преодоления его защиты 19 . Этот эффект Allee создает сильную связь между поведением жуков при отборе деревьев-хозяев и восприимчивостью деревьев-хозяев к колонизации 19,20,21 . Ключевым защитным механизмом хвойных деревьев от нападения жуков-короедов является заливка отверстий жуков смолой, которая физически изгоняет колонизирующих жуков, может быть токсичной для колонизаторов и их грибков и может прерывать общение жуков 22,23 . В нормальных условиях ослабленные деревья с нарушенной защитой наиболее подвержены колонизации и будут основными целями первичных короедов, таких как WPB 9. 0145 13,23,24 . Однако при сильном дефиците воды у многих деревьев больше нет ресурсов для создания защиты 7,13 . Засуха 12,25,26,27 , особенно в сочетании с высокими температурами 24,28,29,30 , может спровоцировать повышенную гибель деревьев, вызванную короедом, по мере снижения средней силы роста деревьев. По мере того, как плотность местной популяции жуков увеличивается из-за успешного размножения в пределах пространственно агрегированных уязвимых деревьев, массовые атаки увеличиваются в размерах и становятся способными сокрушать грозную защиту деревьев. Даже большие здоровые деревья могут быть подвержены колонизации и гибели при высокой плотности популяции жуков 13,23,24 . Таким образом, водный стресс и плотность популяции жуков влияют на восприимчивость отдельных деревьев к жукам-короедам. Когда экстремальная или длительная засуха увеличивает уязвимость дерева-хозяина, темпы роста популяции короеда увеличиваются, а затем становятся самоусиливающимися, поскольку большая плотность жуков делает дополнительные деревья-хозяева уязвимыми для успешного массового нападения 12,13,15,24 .
0145 13,23,24 . Однако при сильном дефиците воды у многих деревьев больше нет ресурсов для создания защиты 7,13 . Засуха 12,25,26,27 , особенно в сочетании с высокими температурами 24,28,29,30 , может спровоцировать повышенную гибель деревьев, вызванную короедом, по мере снижения средней силы роста деревьев. По мере того, как плотность местной популяции жуков увеличивается из-за успешного размножения в пределах пространственно агрегированных уязвимых деревьев, массовые атаки увеличиваются в размерах и становятся способными сокрушать грозную защиту деревьев. Даже большие здоровые деревья могут быть подвержены колонизации и гибели при высокой плотности популяции жуков 13,23,24 . Таким образом, водный стресс и плотность популяции жуков влияют на восприимчивость отдельных деревьев к жукам-короедам. Когда экстремальная или длительная засуха увеличивает уязвимость дерева-хозяина, темпы роста популяции короеда увеличиваются, а затем становятся самоусиливающимися, поскольку большая плотность жуков делает дополнительные деревья-хозяева уязвимыми для успешного массового нападения 12,13,15,24 .
Активность WPB сильно зависит от структуры леса — пространственного расположения и распределения размеров деревьев — и породного состава деревьев. Принимая во внимание только структуру леса, леса с высокой плотностью более подвержены гибели деревьев, вызванной короедом, по сравнению с изреженными лесами 6,9 , что может быть связано с усилением конкуренции за водные ресурсы среди скученных деревьев, что снижает среднюю сопротивляемость деревьев 31 , или с тем, что меньшие промежутки между деревьями защищают шлейфы феромонов от рассеивания ветром и, таким образом, улучшают внутривидовую коммуникацию жуков 32 . Размер дерева является еще одним аспектом структуры леса, который влияет на поведение короеда при выборе хозяина: более мелкие деревья, как правило, имеют меньшую способность противостоять нападению, но более крупные деревья являются более желательными целями из-за их более толстой флоэмы, обеспечивающей большее содержание питательных веществ 13,33,34,35 . Во время вспышки некоторые виды короедов коллективно «переключают» предпочтительный размер дерева для атаки, чтобы найти компромисс между восприимчивостью и качеством хозяина 13,21,36,37,38,39 . Если взять только состав леса, активность WPB в горном хребте Сьерра-Невада в Калифорнии обязательно связана с региональным распространением его исключительного хозяина, сосны пондероза 18 . Колонизация первичными короедами также может зависеть от локальной относительной частоты встречаемости древесных пород в лесных насаждениях, отражая более общую закономерность, заключающуюся в том, что специализированные травоядные насекомые, как правило, ниже в таксономически разнообразных лесах по сравнению с монокультурами 40,41 .
Во время вспышки некоторые виды короедов коллективно «переключают» предпочтительный размер дерева для атаки, чтобы найти компромисс между восприимчивостью и качеством хозяина 13,21,36,37,38,39 . Если взять только состав леса, активность WPB в горном хребте Сьерра-Невада в Калифорнии обязательно связана с региональным распространением его исключительного хозяина, сосны пондероза 18 . Колонизация первичными короедами также может зависеть от локальной относительной частоты встречаемости древесных пород в лесных насаждениях, отражая более общую закономерность, заключающуюся в том, что специализированные травоядные насекомые, как правило, ниже в таксономически разнообразных лесах по сравнению с монокультурами 40,41 .
Взаимодействие между структурой и составом леса как в древостоях, так и в масштабах деревьев также влияет на активность WPB. Например, в густых лесных насаждениях с высокой доступностью хозяев может наблюдаться более высокая смертность деревьев, вызванная жуками, поскольку расстояния расселения между потенциальными деревьями-хозяевами короче, что снижает хищничество взрослых особей, ищущих хозяев, и способствует более высокой скорости колонизации 33,42,43 . Высокая доступность хоста также может снизить вероятность того, что отдельные жуки будут тратить свои ограниченные ресурсы впустую, летя и приземляясь на дерево, не являющееся хозяином 9.0145 44,45 . В более мелком масштабе защитная способность дерева-хозяина может зависеть от положения его кроны со сниженной биохимической защитой у подавленных, скученных деревьев 46 . Таким образом, грубые измерения структуры и состава леса могут лишь частично объяснить механизмы, влияющие на беспокойство короедов. Также необходима более подробная информация, в которой явно указаны виды деревьев, их размер и местная плотность, что лучше отражает экологические процессы, лежащие в основе гибели деревьев, вызванной насекомыми 28,36,38,39 .
Высокая доступность хоста также может снизить вероятность того, что отдельные жуки будут тратить свои ограниченные ресурсы впустую, летя и приземляясь на дерево, не являющееся хозяином 9.0145 44,45 . В более мелком масштабе защитная способность дерева-хозяина может зависеть от положения его кроны со сниженной биохимической защитой у подавленных, скученных деревьев 46 . Таким образом, грубые измерения структуры и состава леса могут лишь частично объяснить механизмы, влияющие на беспокойство короедов. Также необходима более подробная информация, в которой явно указаны виды деревьев, их размер и местная плотность, что лучше отражает экологические процессы, лежащие в основе гибели деревьев, вызванной насекомыми 28,36,38,39 .
Огромные пространственные масштабы гибели деревьев, вызванной WPB, во время жаркой засухи в Калифорнии в 2012–2016 гг. бросают вызов нашей способности одновременно учитывать, как широкомасштабные условия окружающей среды могут взаимодействовать с местной структурой и составом леса, чтобы влиять на динамику между выбором короеда и колонизацией. деревьев-хозяев и восприимчивость деревьев-хозяев к атаке 15,47 . Измерение местной структуры леса обычно требует дорогостоящего оборудования 4,48 или трудоемкие полевые обследования 14,15,49 , что ограничивает объем и частоту обследований. Небольшие беспилотные авиационные системы (БАС) позволяют относительно быстро и дешево получать дистанционные изображения сотен гектаров леса, которые можно использовать для измерения сложной структуры и состава леса в масштабе отдельных деревьев с помощью фотограмметрии «Структура из движения» (SfM) 50,51 . Сверхвысокое сантиметровое разрешение измерений, полученных с помощью sUAS, а также возможность учитывать отражательную способность растительности могут помочь преодолеть проблемы с классификацией видов и обнаружением мертвых деревьев, присущие другим методам дистанционного зондирования, таким как бортовой LiDAR 9.0145 52 . Распределение таких съемок по градиенту окружающей среды может решить проблему сбора данных, присущую исследованию явлений как с сильным локальным, так и с сильным широкомасштабным компонентом.
деревьев-хозяев и восприимчивость деревьев-хозяев к атаке 15,47 . Измерение местной структуры леса обычно требует дорогостоящего оборудования 4,48 или трудоемкие полевые обследования 14,15,49 , что ограничивает объем и частоту обследований. Небольшие беспилотные авиационные системы (БАС) позволяют относительно быстро и дешево получать дистанционные изображения сотен гектаров леса, которые можно использовать для измерения сложной структуры и состава леса в масштабе отдельных деревьев с помощью фотограмметрии «Структура из движения» (SfM) 50,51 . Сверхвысокое сантиметровое разрешение измерений, полученных с помощью sUAS, а также возможность учитывать отражательную способность растительности могут помочь преодолеть проблемы с классификацией видов и обнаружением мертвых деревьев, присущие другим методам дистанционного зондирования, таким как бортовой LiDAR 9.0145 52 . Распределение таких съемок по градиенту окружающей среды может решить проблему сбора данных, присущую исследованию явлений как с сильным локальным, так и с сильным широкомасштабным компонентом.
Мы использовали изображения дистанционного зондирования, полученные с помощью sUAS, на сети из 32 участков в сосновых/смешанных хвойных лесах Сьерра-Невада пондероза, охватывающих 1000 м над уровнем моря и 350 км широты 14 , охватывающих в общей сложности 9 км 2 , изучить, как широкомасштабные условия окружающей среды взаимодействовали с местной структурой и составом леса, чтобы сформировать закономерности гибели деревьев во время кумулятивного случая гибели деревьев с 2012 по 2018 год. Мы спросили:
- 1.
Как пропорция деревьев-хозяев пондерозовой сосны в данной местности и средний размер деревьев-хозяев влияют на гибель деревьев, вызванную WPB?
- 2.
Как плотность всех деревьев (далее «общая плотность») влияет на гибель деревьев, вызванную WPB?
- 3.
Как общая базовая площадь всех деревьев (далее «общая базальная площадь») влияет на гибель деревьев, вызванную WPB?
- 4.
Как вызванный окружающей средой стресс от влажности деревьев влияет на гибель деревьев, вызванную WPB?
- 5.
Как влияют структура леса, состав леса и условия окружающей среды на гибель деревьев, вызванную WPB?

Здесь мы показываем, что большая локальная доля деревьев-хозяев (сосна пондероза) значительно увеличивает вероятность гибели хозяев, а большая плотность хозяев усиливает этот эффект. Мы также показываем, что более высокий уровень CWD на уровне сайта увеличивает смертность хозяев. Кроме того, мы показываем, что более крупные деревья-хозяева увеличивают вероятность гибели хозяев в соответствии с хорошо известной историей жизни WPB. Важно отметить, что мы обнаруживаем сильную взаимосвязь между размером хоста и CWD, так что уровень смертности хостов особенно высок на жарких/сухих участках, где локальный средний размер дерева хостов велик. Наши результаты демонстрируют межмасштабное взаимодействие в реакции WPB на локальную структуру и состав леса в зависимости от градиента окружающей среды, что помогает согласовать различия между наблюдаемыми моделями гибели деревьев в масштабах всей экосистемы и прогнозами моделей, основанных на более крупномасштабной структуре леса.
Важно отметить, что мы обнаруживаем сильную взаимосвязь между размером хоста и CWD, так что уровень смертности хостов особенно высок на жарких/сухих участках, где локальный средний размер дерева хостов велик. Наши результаты демонстрируют межмасштабное взаимодействие в реакции WPB на локальную структуру и состав леса в зависимости от градиента окружающей среды, что помогает согласовать различия между наблюдаемыми моделями гибели деревьев в масштабах всей экосистемы и прогнозами моделей, основанных на более крупномасштабной структуре леса.
Результаты
Производительность алгоритма обнаружения деревьев
Мы обнаружили, что экспериментальный алгоритм lmfx 53 со значениями параметров dist2d = 1 и ws = 2,5 показал лучшие результаты по семи показателям структуры леса, измеренным с помощью корреляции Пирсона с наземными данными ( Таблица 1).
Таблица 1 Корреляция и различия между наиболее эффективным алгоритмом обнаружения деревьев (lmfx с dist2d = 1 и ws = 2,5) и наземными данными.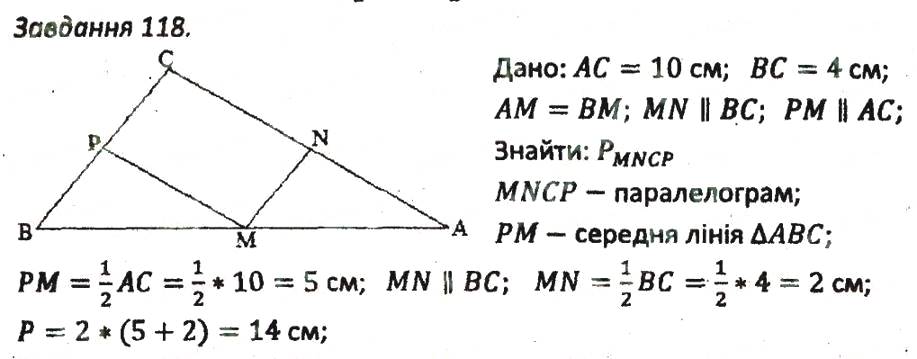
Полноразмерный стол
Точность классификации для живого/мертвого и хозяина/не хозяина
Точность классификации живого/мертвого на закрытом наборе тестовых данных составила 96,4%. Точность классификации видов по скрытому набору данных тестирования составила 64,1%. Точность хозяина WPB/не-хозяина WPB (т. е. сосна пондероза по сравнению с другими породами деревьев) в скрытом наборе данных тестирования составила 71,8%.
Сводка сайта на основе лучшего алгоритма обнаружения деревьев и классификации
На всех исследуемых участках мы обнаружили, сегментировали и классифицировали 452 413 деревьев в 23 187 пикселях размером 20 × 20 м (при этом площадь каждого пикселя приблизительно эквивалентна площади полевой участок). Из этих деревьев мы классифицировали 118 879.как умершие (смертность 26,3%). Оценочная смертность деревьев на уровне участка колебалась от 6,8% до 53,6%. См. дополнительную таблицу 1, где приведены сводные данные по участкам и сравнения со смертностью на уровне участков, измеренной по полевым данным.
Влияние местной структуры и регионального климата на смертность деревьев, приписываемую WPB
CWD на уровне участка оказал положительное основное влияние на вероятность гибели пондерозы (величина эффекта: 0,85; 95% ДИ: [0,70, 0,99]; рис. 1). Мы обнаружили положительный основной эффект доли деревьев-хозяев на клетку (величина эффекта: 0,68; 95% ДИ: [0,62, 0,74]), с большей долей деревьев-хозяев (например, сосны желтой) в клетке, увеличивающей вероятность гибели сосны желтой. Мы не обнаружили влияния общей плотности деревьев или общей площади основания (т. е. включая как сосну пондерозу, так и виды, не являющиеся хозяевами; размер эффекта плотности деревьев: -0,01; 95% ДИ: [-0,11, 0,08]; размер эффекта площади основания: — 0,13; 95% ДИ: [-0,29, 0,03]).
Рис. 1: Апостериорные распределения величины эффекта из биномиальной модели с нулевым раздуванием, предсказывающей вероятность гибели сосны желтой в ячейке 20 × 20 м с учетом характеристик структуры леса и климатического дефицита воды (КВД) на уровне участка.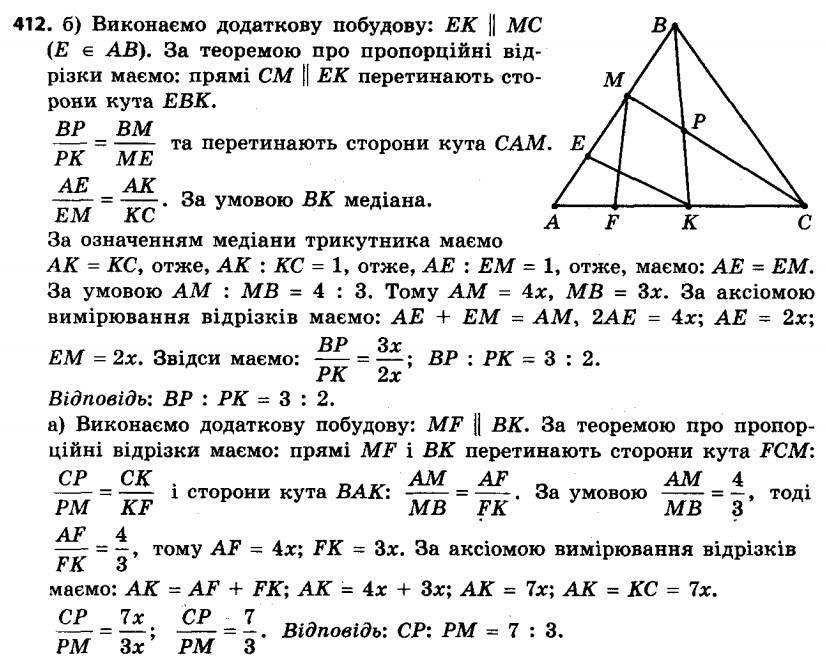
Закрашенная серым цветом область для каждой ковариаты модели представляет плотность вероятности апостериорного распределения, точка под каждой кривой плотности представляет собой медиану оценки, жирный интервал вокруг точечной оценки представляет 66-процентный достоверный интервал, а тонкий интервал вокруг точечной оценки представляет собой 95% достоверный интервал. Оценки всех параметров модели, включая параметры гауссовского процесса для каждого участка, можно найти в дополнительной таблице 2.
Изображение полного размера
Мы обнаружили положительное двустороннее взаимодействие между общей плотностью деревьев на ячейку и долей деревьев-хозяев, что эквивалентно положительному влиянию плотности деревьев-хозяев (величина эффекта: 0,06; 95% ДИ: [0,01, 0,12]; рис. 1).
Мы обнаружили положительное основное влияние средней высоты сосны пондероза на вероятность гибели пондерозы (величина эффекта: 0,25; 95% ДИ: [0,14, 0,35]). В сочетании с сильной корреляцией между долей мертвых деревьев-хозяев и убитой базальной площадью (дополнительный рисунок 1 и дополнительное примечание 1) эти результаты показывают, что WPB в среднем атаковал более крупные деревья. Кроме того, наблюдалась сильная положительная взаимосвязь между CWD и средней высотой сосны пондерозы, так что более крупные деревья с особой вероятностью увеличивали локальную вероятность гибели пондерозы в более жарких и засушливых местах (величина эффекта: 0,54; 9).5% ДИ: [0,37, 0,70]; Рис. 2).
Кроме того, наблюдалась сильная положительная взаимосвязь между CWD и средней высотой сосны пондерозы, так что более крупные деревья с особой вероятностью увеличивали локальную вероятность гибели пондерозы в более жарких и засушливых местах (величина эффекта: 0,54; 9).5% ДИ: [0,37, 0,70]; Рис. 2).
Линия «более крупные деревья» представляет собой среднюю высоту сосны пондерозовой на 0,7 стандартных отклонения выше среднего (приблизительно 24,1 м), а линия «более мелкие деревья» представляет среднюю высоту сосны пондерозовой на 0,7 стандартных отклонения ниже среднего (~12,1 м). м).
Полноразмерное изображение
Мы не обнаружили влияния взаимодействий CWD на уровне сайта с долей деревьев-хозяев (величина эффекта: −0,08; 95% ДИ: [−0,18, 0,03]) или взаимодействия между CWD и общим базальная площадь (величина эффекта: -0,04; 95% ДИ: [-0,23, 0,15]; рис. 1).
1).
Мы обнаружили отрицательный эффект взаимодействия CWD с общей плотностью деревьев (величина эффекта: -0,19; 95% ДИ: [-0,31, -0,07]), а также взаимодействия между средней высотой деревьев-хозяев и общей базальная площадь (величина эффекта: −0,08; 95% ДИ: [-0,13, -0,03]; Рисунок 1).
Хотя мы не обнаружили взаимодействия между долей деревьев-хозяев и средней высотой дерева-хозяина, мы обнаружили трехстороннее взаимодействие между этими переменными с CWD (величина эффекта: 0,14; 95% ДИ: [0,04, 0,24]; рис. 1).
Обсуждение
В этом исследовании используются изображения, полученные с помощью дронов, чтобы уточнить наше понимание закономерностей гибели деревьев после жаркой засухи в Калифорнии 2012–2016 годов и ее последствий. Одновременно измеряя влияние местной структуры и состава леса на широкомасштабные градиенты окружающей среды, мы смогли лучше охарактеризовать влияние насекомого, убивающего деревья, WPB, по сравнению с использованием только коррелятов стресса деревьев.
Сильный положительный основной эффект CWD
Мы обнаружили сильное положительное влияние CWD на уровне участка на смертность сосны желтой. Мы не измеряли водный стресс деревьев на уровне отдельных деревьев, как в другой недавней работе 15 , а вместо этого рассматривали CWD как общий показатель стресса деревьев по результатам более грубых исследований 11 . При измерении в мелком масштабе, даже если не на уровне отдельного дерева, прогрессирующая потеря воды в кронах деревьев может быть хорошим индикатором водного стресса деревьев и повышенной уязвимости к гибели от засухи или короедов 5 . Несмотря на то, что в период с 2012 по 2016 год на всей исследуемой территории наблюдалась исключительная жаркая засуха 2,3 , использование среднего значения CWD за 30 лет в качестве индикатора стресса деревьев на уровне участка не позволяет нам понять, была ли доступность воды ниже в абсолютный смысл во время засухи или повышение уязвимости деревьев перед короедом было вызвано хроническим дефицитом воды в этих исторически более жарких/сухих местах 54 .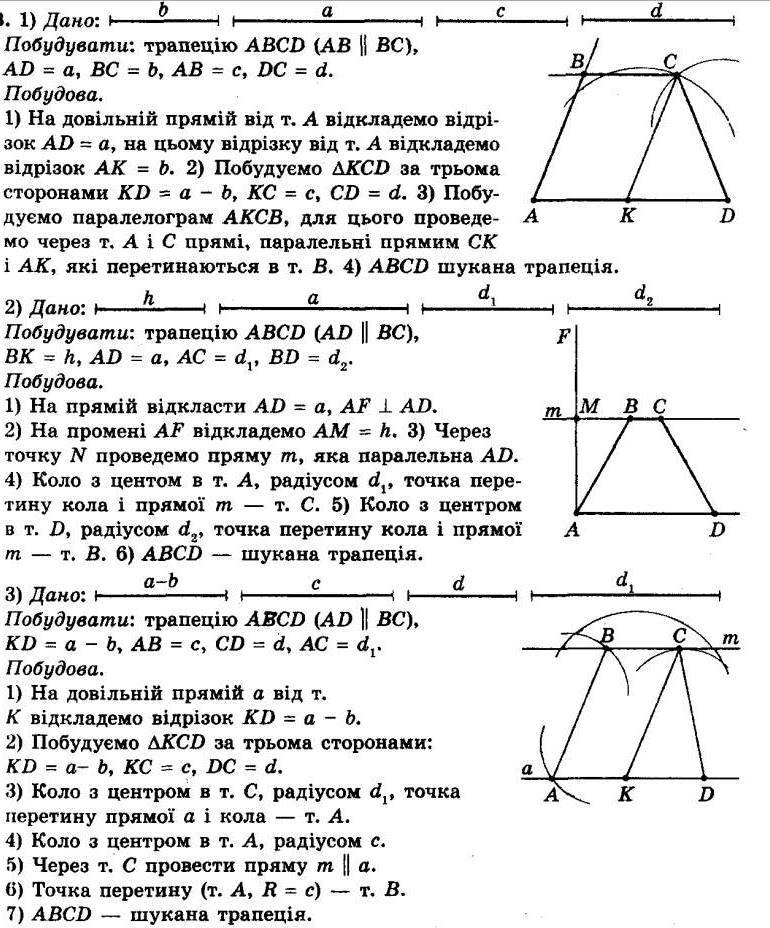
Положительный эффект пропорции и плотности хозяина
Ряд механизмов, связанных с относительной численностью видов в данной местности, может лежать в основе сильного влияния доли хозяев на вероятность гибели деревьев-хозяев. Зависимая от частоты травоядность — при этом смешанные леса менее травоядны по сравнению с монокультурами (в качестве крайнего примера) — распространена, особенно для видов насекомых-олигофагов 40 . Летучие вещества, не являющиеся хозяином, снижают привлекательность некоторых видов короедов к их агрегационным феромонам 55 , в том числе WPB 56 . Комбинации летучих веществ, не являющихся хозяевами, и феромона, препятствующего агрегации, успешно использовались для снижения уровня смертности деревьев, связанной с WPB, в Калифорнии 57,58 . Положительная взаимосвязь между плотностью растений-хозяев и восприимчивостью к колонизации жуками-короедами была настолько хорошо задокументирована на уровне экспериментального участка 43,59,60 , что снижение плотности насаждений за счет селективного сбора растений-хозяев обычно рекомендуется для снижения уровня древесной растительности в будущем. смертность, приписываемая короедам 61 , в том числе WPB 18 . Большая плотность растений-хозяев сокращает расстояние полета, необходимое WPB для распространения на новых хозяев, что, вероятно, способствует распространению короедов, однако мы откалибровали наше воздушное обнаружение деревьев на ~400 м 2 площадей, а не на отдельные деревья, поэтому наши данные недостаточны для разрешения этих отношений. Повышенная плотность сосны пондероза, в частности, может непропорционально увеличить конкурентную среду для деревьев-хозяев (и, таким образом, повысить их восприимчивость к колонизации WPB), если внутривидовая конкуренция среди деревьев сосны пондероза сильнее, чем межвидовая конкуренция, как предсказывает теория сосуществования 9.0145 62 . Наконец, более высокая плотность хозяев увеличивает частоту, когда поиск WPB приземляется на хозяев, а не на не-хозяев, тем самым уменьшая количество энергии, затрачиваемой на поиск и выбор хозяина, а также время, которое поиск WPB тратит на воздействие различных хищников за пределами дома.
смертность, приписываемая короедам 61 , в том числе WPB 18 . Большая плотность растений-хозяев сокращает расстояние полета, необходимое WPB для распространения на новых хозяев, что, вероятно, способствует распространению короедов, однако мы откалибровали наше воздушное обнаружение деревьев на ~400 м 2 площадей, а не на отдельные деревья, поэтому наши данные недостаточны для разрешения этих отношений. Повышенная плотность сосны пондероза, в частности, может непропорционально увеличить конкурентную среду для деревьев-хозяев (и, таким образом, повысить их восприимчивость к колонизации WPB), если внутривидовая конкуренция среди деревьев сосны пондероза сильнее, чем межвидовая конкуренция, как предсказывает теория сосуществования 9.0145 62 . Наконец, более высокая плотность хозяев увеличивает частоту, когда поиск WPB приземляется на хозяев, а не на не-хозяев, тем самым уменьшая количество энергии, затрачиваемой на поиск и выбор хозяина, а также время, которое поиск WPB тратит на воздействие различных хищников за пределами дома. хост-дерево.
хост-дерево.
Нет основного влияния общей плотности, но есть взаимодействие с CWD
Мы не обнаружили связи между общей плотностью деревьев и гибелью сосны желтой, хотя работа на совпадающих наземных участках показала отрицательную, но слабую связь при использовании доли деревьев, погибших в качестве ответа 14 . Кайзер и др. 28 также показывают заражение большим горным сосновым жуком ( Dendroctonus ponderosae ) на участках с низкой плотностью населения в Монтане. Однако Hayes et al. 31 и Fettig et al. 14 обнаружили, что показатели общей плотности деревьев объясняют большую изменчивость смертности деревьев, чем показатели доступности растений-хозяев, хотя эти выводы были основаны на более широком анализе 31 или на другой переменной ответа (например, «общее количество мертвых деревьев-хозяев 14 , а не биномиальный ответ «количество мертвых деревьев-хозяев зависит от общего количества деревьев-хозяев», как в нашем исследовании).
Больший размер нашей выборки, возможно, позволил нам более точно проанализировать роль многогранной структуры и состава леса, а также CWD и взаимодействий в факторах смертности пондерозовой сосны. Действительно, мы обнаружили отрицательное двустороннее взаимодействие между CWD участка и общей плотностью, предполагая, что в более густых насаждениях наблюдался более низкий уровень смертности пондерозы в более жарких и засушливых местах, что согласуется с Restaino et al. 9 в результате их неконтролируемого градиента общей плотности в том же регионе во время той же жаркой засухи. В отсутствие активного управления структура леса в значительной степени является продуктом климата и, что приобретает все большее значение в более мелких пространственных масштабах, топографических условий 63 . Более густые участки леса в нашем исследовании могут указывать на большую местную доступность воды, более благоприятные условия для роста и выживания деревьев, а также на повышенную устойчивость к гибели деревьев, вызванной жуками, особенно когда более густые участки находятся в жарких и сухих местах 9,63,64 .
Влияние общей площади основания
В то время как общая плотность деревьев, вероятно, является индикатором благоприятных микроучастков в пожаротушенных лесах, общая площадь основания является лучшим показателем местной конкурентной среды, особенно в лесах с ограниченным водоснабжением 63,64 . Однако мы не обнаружили основного влияния общей базальной площади на вероятность гибели пондерозы, а также ее взаимодействия с CWD на уровне участка. Это контрастирует с результатами Young et al. 11 и из анализа совпадающих полевых участков 14 . В отличие от Young et al. 11 может быть объяснено разными масштабами анализа (например, 3500 × 3500 пикселей против 20 × 20), контраст с совпадающими наземными графиками более загадочен. Одно из объяснений заключается в том, что отбор проб дронами захватил большую площадь за пределами условно отобранных полевых участков (т. е. критерием выбора участка было 10% гибели базальной площади сосны пондерозы), что отражало другую взаимосвязь между местной базальной площадью и смертностью деревьев. Возможно, более вероятно, что наше измерение общей базальной площади недостаточно точно для представления местной конкурентной среды по сравнению с базальной площадью, полученной в полевых условиях. Для нашего исследования базальная площадь была получена из видоспецифичных и изначально зашумленных аллометрических взаимосвязей с высотой дерева, которая сама была получена в результате обработки изображений с дрона SfM. По мере совершенствования технологии дистанционного зондирования, позволяющей извлекать более точную информацию (например, измерения отдельных деревьев), расширяется диалог между экологами всех мастей 65,66,67 необходим, чтобы полностью представить, как лучше всего дистанционно измерять природные явления, либо используя уже изобретенные колеса (например, базовую площадь участка), либо вводя что-то совершенно новое.
Возможно, более вероятно, что наше измерение общей базальной площади недостаточно точно для представления местной конкурентной среды по сравнению с базальной площадью, полученной в полевых условиях. Для нашего исследования базальная площадь была получена из видоспецифичных и изначально зашумленных аллометрических взаимосвязей с высотой дерева, которая сама была получена в результате обработки изображений с дрона SfM. По мере совершенствования технологии дистанционного зондирования, позволяющей извлекать более точную информацию (например, измерения отдельных деревьев), расширяется диалог между экологами всех мастей 65,66,67 необходим, чтобы полностью представить, как лучше всего дистанционно измерять природные явления, либо используя уже изобретенные колеса (например, базовую площадь участка), либо вводя что-то совершенно новое.
Положительный основной эффект среднего размера дерева-хозяина
Положительный основной эффект среднего размера дерева-хозяина на уровень смертности пондероза соответствует общепринятому мнению о динамике WPB в Сьерра-Неваде, а также других первичных короедов 18 . WPB отдает предпочтение деревьям 50,8–76,2 см DBH 9Fettig et al. 14 на совпадающих полевых участках, а также в других недавних исследованиях 9,15,70 . Большие деревья более питательны и поэтому являются идеальными целями, если местная плотность жуков-короедов достаточно высока, чтобы успешно инициировать массовое нападение и сокрушить защиту деревьев, что может произойти, когда многие деревья находятся в условиях водного стресса 7,13,24 . Во время недавней жаркой засухи мы ожидали, что большинство деревьев будут испытывать сильный водный стресс, что подготовило почву для увеличения плотности жуков, успешных массовых атак и нападения на более крупные деревья. Учитывая, что наша калибровка высоты мертвого дерева была консервативной (с учетом недооценки высоты мертвого дерева, полученной с помощью дронов, по сравнению с деревьями, измеренными в полевых условиях), вполне вероятно, что положительный основной эффект высоты дерева, о котором мы сообщаем, представляет собой нижнюю границу этого эффекта.
WPB отдает предпочтение деревьям 50,8–76,2 см DBH 9Fettig et al. 14 на совпадающих полевых участках, а также в других недавних исследованиях 9,15,70 . Большие деревья более питательны и поэтому являются идеальными целями, если местная плотность жуков-короедов достаточно высока, чтобы успешно инициировать массовое нападение и сокрушить защиту деревьев, что может произойти, когда многие деревья находятся в условиях водного стресса 7,13,24 . Во время недавней жаркой засухи мы ожидали, что большинство деревьев будут испытывать сильный водный стресс, что подготовило почву для увеличения плотности жуков, успешных массовых атак и нападения на более крупные деревья. Учитывая, что наша калибровка высоты мертвого дерева была консервативной (с учетом недооценки высоты мертвого дерева, полученной с помощью дронов, по сравнению с деревьями, измеренными в полевых условиях), вполне вероятно, что положительный основной эффект высоты дерева, о котором мы сообщаем, представляет собой нижнюю границу этого эффекта.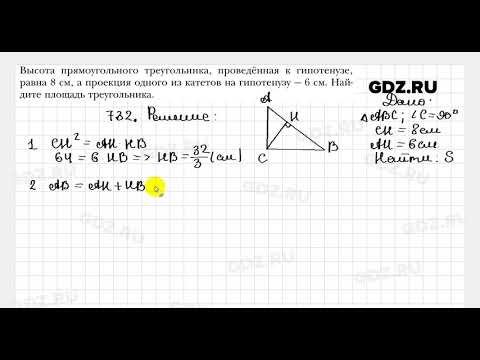 Кроме того, Феттиг и соавт. 14 не обнаружил связи между размером дерева и смертностью для ладана или белой пихты на совпадающих полевых участках. Эти виды составляют 22,3% от общей смертности деревьев, наблюдаемой в их исследовании, однако в нашем исследовании все мертвые деревья были классифицированы как сосна пондероза (см. раздел «Методы»), что могло еще больше ослабить положительное влияние размера дерева на смертность деревьев. что мы идентифицировали.
Кроме того, Феттиг и соавт. 14 не обнаружил связи между размером дерева и смертностью для ладана или белой пихты на совпадающих полевых участках. Эти виды составляют 22,3% от общей смертности деревьев, наблюдаемой в их исследовании, однако в нашем исследовании все мертвые деревья были классифицированы как сосна пондероза (см. раздел «Методы»), что могло еще больше ослабить положительное влияние размера дерева на смертность деревьев. что мы идентифицировали.
Межмасштабное взаимодействие CWD и размера дерева-хозяина
В более жарких и засушливых местах больший средний размер хозяина увеличивает вероятность его гибели. Примечательно, что аналогичная картина была показана Stovall et al. 65 в исследовании, проведенном в южной части Сьерра-Невады (т. е. в самой жаркой и засушливой части более пространственно обширных результатов, которые мы представляем здесь) с сильной положительной зависимостью высоты деревьев от смертности в районах с наибольшим дефицитом давления пара и отсутствием деревьев. соотношение роста и смертности в районах с наименьшим дефицитом давления пара. Наша работа предполагает, что WPB ориентировался на различные аспекты структуры леса в зависимости от градиента окружающей среды в пространственном контексте параллельно с временным контекстом, отмеченным Stovall et al. 65 и Pile et al. 70 , которые заметили, что смертность все больше была вызвана более крупными деревьями по мере того, как жаркая засуха продолжалась и становилась все более сильной. Временной сигнал о том, что жуки-короеды нападают на все более и более крупные деревья-хозяева, отражает положительную обратную связь между структурой леса и динамикой популяции жуков-короедов по мере смены фазы популяции от эндемичной к эпидемической 13 . Эта положительная обратная связь, приводящая к эруптивной динамике популяции, хорошо задокументирована как временное явление, и здесь мы показываем аналогичную картину в пространственном контексте, опосредованную CWD на уровне сайта.
соотношение роста и смертности в районах с наименьшим дефицитом давления пара. Наша работа предполагает, что WPB ориентировался на различные аспекты структуры леса в зависимости от градиента окружающей среды в пространственном контексте параллельно с временным контекстом, отмеченным Stovall et al. 65 и Pile et al. 70 , которые заметили, что смертность все больше была вызвана более крупными деревьями по мере того, как жаркая засуха продолжалась и становилась все более сильной. Временной сигнал о том, что жуки-короеды нападают на все более и более крупные деревья-хозяева, отражает положительную обратную связь между структурой леса и динамикой популяции жуков-короедов по мере смены фазы популяции от эндемичной к эпидемической 13 . Эта положительная обратная связь, приводящая к эруптивной динамике популяции, хорошо задокументирована как временное явление, и здесь мы показываем аналогичную картину в пространственном контексте, опосредованную CWD на уровне сайта.
Ключевое отличие от положительной обратной связи от эндемии к эпидемии, отмеченной Boone et al. 13 заключается в том, что ни один из районов нашего исследования не считался находящимся в эндемичной популяционной фазе по типичным показателям динамики WPB 31,33 . Динамика WPB на всех участках считалась эпидемической: погибло более 5 деревьев на гектар (дополнительная таблица 1). Межмасштабное взаимодействие между широкомасштабным CWD и размером дерева-хозяина в локальном масштабе, даже среди популяций, находящихся в фазе эпидемии, подчеркивает драматические последствия положительной обратной связи для смертности деревьев в ландшафтном масштабе. Массовая гибель деревьев в более жарких/сухих лесах Сьерра-Невады (более низкие широты и высоты 4,11 ) в период с 2012 по 2016 год жаркая засуха, вероятно, возникла как синергетическое выравнивание условий окружающей среды и местной структуры леса, что позволило WPB успешно заселить большие деревья, быстро увеличить размер популяции и расшириться. Неожиданно низкая смертность в более прохладных/влажных лесах Сьерра-Невады по сравнению с прогнозами модели, основанными на более крупномасштабных данных о структуре леса 11 , может быть результатом различной реакции WPB на местную структуру леса из-за отсутствия согласования с благоприятными климатическими условиями и слабые положительные отзывы.
Неожиданно низкая смертность в более прохладных/влажных лесах Сьерра-Невады по сравнению с прогнозами модели, основанными на более крупномасштабных данных о структуре леса 11 , может быть результатом различной реакции WPB на местную структуру леса из-за отсутствия согласования с благоприятными климатическими условиями и слабые положительные отзывы.
Ограничения и будущие направления
Мы продемонстрировали, что дроны могут быть эффективным средством сбора данных о лесах в различных, совершенно разных пространственных масштабах для исследования одного многомасштабного явления — от нескольких метров между деревьями до сотен метров над уровнем моря. , до сотен тысяч метров широты. Некоторые ограничения остаются, но их можно преодолеть путем усовершенствования использования этого инструмента для экологии леса. Большинство этих ограничений связано с классификацией и измерением стоящих мертвых деревьев, поэтому необходимо работать с полевыми данными для калибровки и составления отчетов о неопределенности.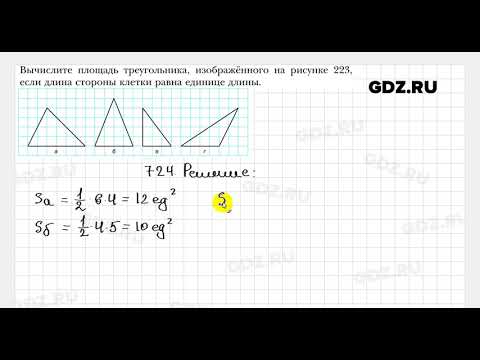
Наибольшее ограничение в нашем исследовании, возникающее из-за неопределенности классификации, заключается в предположении, что все мертвые деревья были сосной пондерозой, которое, по нашим оценкам на совпадающих полевых участках, верно примерно в 73,4% случаев. Поскольку факторы структуры леса, влияющие на вероятность гибели отдельных деревьев во время жаркой засухи, зависели от древесных пород 15 , мы не можем исключить, что некоторые наблюдаемые нами взаимосвязи смертности сосны желтоватой со структурой леса могут быть частично объяснены этими взаимосвязями в другие виды, которые были ошибочно классифицированы как сосна пондероза с использованием наших методов. Тем не менее, общий состав сообщества в районе нашего исследования был схожим 14 , и мы можем воспроизвести аналогичные модели структуры леса / смертности в данных, полученных с помощью дронов, если ограничить объем анализа только деревьями, обнаруженными в следах на совпадающих полевых участках (дополнительный рисунок 2). Таким образом, мы по-прежнему уверены, что наблюдаемые нами закономерности были обусловлены в первую очередь динамикой между WPB и сосной пондероза. В то время как спектральная информация о листве могла помочь классифицировать живые деревья по видам, виды стоящих мертвых деревьев не были спектрально различимы. Эта задача классификации стоячих мертвых деревьев по видам подразумевает, что системы хвойных лесов с меньшим разнообразием короедов и деревьев-хозяев, такие как вспышки горного соснового лубоеда в относительных монокультурах естественных сосновых лесов скрученной сосны в Межгорном Западе, должны быть особенно поддающимися воздействию методы, представленные здесь, даже с минимальными дальнейшими уточнениями, потому что мертвые деревья почти наверняка будут принадлежать одному виду и подверглись колонизации одним видом жуков-короедов. По тем же причинам эти методы также будут работать особенно хорошо, если изображения будут сняты до события смерти.
Таким образом, мы по-прежнему уверены, что наблюдаемые нами закономерности были обусловлены в первую очередь динамикой между WPB и сосной пондероза. В то время как спектральная информация о листве могла помочь классифицировать живые деревья по видам, виды стоящих мертвых деревьев не были спектрально различимы. Эта задача классификации стоячих мертвых деревьев по видам подразумевает, что системы хвойных лесов с меньшим разнообразием короедов и деревьев-хозяев, такие как вспышки горного соснового лубоеда в относительных монокультурах естественных сосновых лесов скрученной сосны в Межгорном Западе, должны быть особенно поддающимися воздействию методы, представленные здесь, даже с минимальными дальнейшими уточнениями, потому что мертвые деревья почти наверняка будут принадлежать одному виду и подверглись колонизации одним видом жуков-короедов. По тем же причинам эти методы также будут работать особенно хорошо, если изображения будут сняты до события смерти.
Некоторая неопределенность окружала нашу способность обнаруживать деревья, используя геометрию плотных облаков точек, полученную с помощью SfM. Горизонтальная точность (т. е. положение долготы/широты) обнаружения дерева была лучше, чем вертикальная точность (т. е. высота), что может быть результатом более значительного вклада ошибок в полевые расчеты высоты дерева по сравнению с относительным положением дерева. до центра графика (таблица 1). Измерения высоты были особенно сложными для стоячих мертвых деревьев, потому что SfM может не дать никаких точек, представляющих узкие вершины деревьев без иголок в результирующем плотном облаке точек. Наша консервативная калибровка высоты дерева, измеренной дронами, до высоты, измеренной в полевых условиях, усилила основное влияние CWD на смертность хозяина в нашей модели и обратила влияние высоты дерева-хозяина. Мы сообщаем, что более крупные деревья-хозяева увеличивают вероятность гибели деревьев-хозяев, в то время как модели, использующие некалиброванные высоты деревьев, показывают, что более крупные деревья снижают уровень смертности хозяев (см. Дополнительный рис. 3 по сравнению с рис.
Горизонтальная точность (т. е. положение долготы/широты) обнаружения дерева была лучше, чем вертикальная точность (т. е. высота), что может быть результатом более значительного вклада ошибок в полевые расчеты высоты дерева по сравнению с относительным положением дерева. до центра графика (таблица 1). Измерения высоты были особенно сложными для стоячих мертвых деревьев, потому что SfM может не дать никаких точек, представляющих узкие вершины деревьев без иголок в результирующем плотном облаке точек. Наша консервативная калибровка высоты дерева, измеренной дронами, до высоты, измеренной в полевых условиях, усилила основное влияние CWD на смертность хозяина в нашей модели и обратила влияние высоты дерева-хозяина. Мы сообщаем, что более крупные деревья-хозяева увеличивают вероятность гибели деревьев-хозяев, в то время как модели, использующие некалиброванные высоты деревьев, показывают, что более крупные деревья снижают уровень смертности хозяев (см. Дополнительный рис. 3 по сравнению с рис. 1). Хотя наша классификация живых/мертвых была довольно точной (96,4% в скрытом наборе данных), наш классификатор видов, вероятно, выиграет от лучшей сегментации кроны, поскольку значения отражательной способности на уровне пикселей в каждой кроне усредняются для характеристики «спектральной сигнатуры» каждого дерева. При лучшем описании кроны каждого дерева среднее значение пикселей в кроне каждого дерева, вероятно, будет более репрезентативным для спектральной характеристики этого дерева.
1). Хотя наша классификация живых/мертвых была довольно точной (96,4% в скрытом наборе данных), наш классификатор видов, вероятно, выиграет от лучшей сегментации кроны, поскольку значения отражательной способности на уровне пикселей в каждой кроне усредняются для характеристики «спектральной сигнатуры» каждого дерева. При лучшем описании кроны каждого дерева среднее значение пикселей в кроне каждого дерева, вероятно, будет более репрезентативным для спектральной характеристики этого дерева.
Лучшее обнаружение деревьев, сегментация кроны и измерение высоты мертвых деревьев, вероятно, улучшатся с помощью более качественных облаков точек SfM, которые можно улучшить за счет большего перекрытия между изображениями 71 или с изображением под углом (т. е. вне надира) 72 . Фрей и др. 71 обнаружили, что 95-процентное перекрытие предпочтительнее для создания плотных облаков точек в лесных районах, а Джеймс и Робсон 72 уменьшили ошибки плотных облаков точек, используя изображения, сделанные под углом 30 градусов от надира.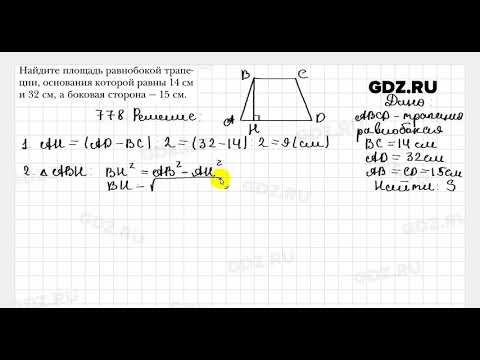 Мы достигли только 91,6% перекрытия с камерой X3 RGB и 83,9% перекрытия с мультиспектральной камерой, и все изображения были обращены к надиру. Мы ожидаем, что компьютерное зрение и глубокое обучение также окажутся полезными в преодолении некоторых из этих проблем обнаружения и классификации9.0145 73 .
Мы достигли только 91,6% перекрытия с камерой X3 RGB и 83,9% перекрытия с мультиспектральной камерой, и все изображения были обращены к надиру. Мы ожидаем, что компьютерное зрение и глубокое обучение также окажутся полезными в преодолении некоторых из этих проблем обнаружения и классификации9.0145 73 .
Наконец, мы отмечаем, что наше исследование ограничено неопределенностью в измерении базальной площади при обработке SfM изображений, полученных с дронов. Эта неопределенность затрудняет представление типичных полевых показателей локальной конкурентной среды (например, общая базальная площадь участка) или воздействия на экосистему (например, доля мертвой базальной площади на участке) в статистическом анализе. Вместо этого мы решили использовать вероятность гибели пондерозы в качестве нашей ключевой переменной ответа, которая хорошо подходит для понимания динамики между поведением колонизации WPB и восприимчивостью дерева-хозяина.
Выводы
Стратегии адаптации к изменению климата делают акцент на управленческих действиях, учитывающих реакцию всей экосистемы на неизбежные изменения 74 , что требует макроэкологического понимания того, как могут взаимодействовать явления в различных масштабах. Уязвимость деревьев к стрессовым факторам окружающей среды представляет собой лишь частичное объяснение моделей гибели деревьев во время жарких засух, особенно при наличии жуков-короедов. Мы показали, что дроны могут быть ценным инструментом для исследования мультискалярных явлений, таких как то, как местная структура леса сочетается с условиями окружающей среды, формируя беспокойство лесных насекомых. Понимание условий, которые определяют реакцию засушливых западных лесов США на нарушения, такие как нашествия короедов, будет иметь жизненно важное значение для прогнозирования последствий увеличения частоты и интенсивности нарушений, усугубляемых изменением климата 75 . Наше исследование предполагает, что результаты будут зависеть от взаимодействия между локальной структурой леса и широкомасштабными экологическими градиентами, при этом межмасштабные взаимодействия могут улучшить наше понимание динамики лесных насекомых.
Уязвимость деревьев к стрессовым факторам окружающей среды представляет собой лишь частичное объяснение моделей гибели деревьев во время жарких засух, особенно при наличии жуков-короедов. Мы показали, что дроны могут быть ценным инструментом для исследования мультискалярных явлений, таких как то, как местная структура леса сочетается с условиями окружающей среды, формируя беспокойство лесных насекомых. Понимание условий, которые определяют реакцию засушливых западных лесов США на нарушения, такие как нашествия короедов, будет иметь жизненно важное значение для прогнозирования последствий увеличения частоты и интенсивности нарушений, усугубляемых изменением климата 75 . Наше исследование предполагает, что результаты будут зависеть от взаимодействия между локальной структурой леса и широкомасштабными экологическими градиентами, при этом межмасштабные взаимодействия могут улучшить наше понимание динамики лесных насекомых.
Методы
Система исследования
Мы разработали аэрофотосъемку так, чтобы она совпадала со 160 участками мониторинга растительности/лесных насекомых на 32 участках, созданных Fettig et al.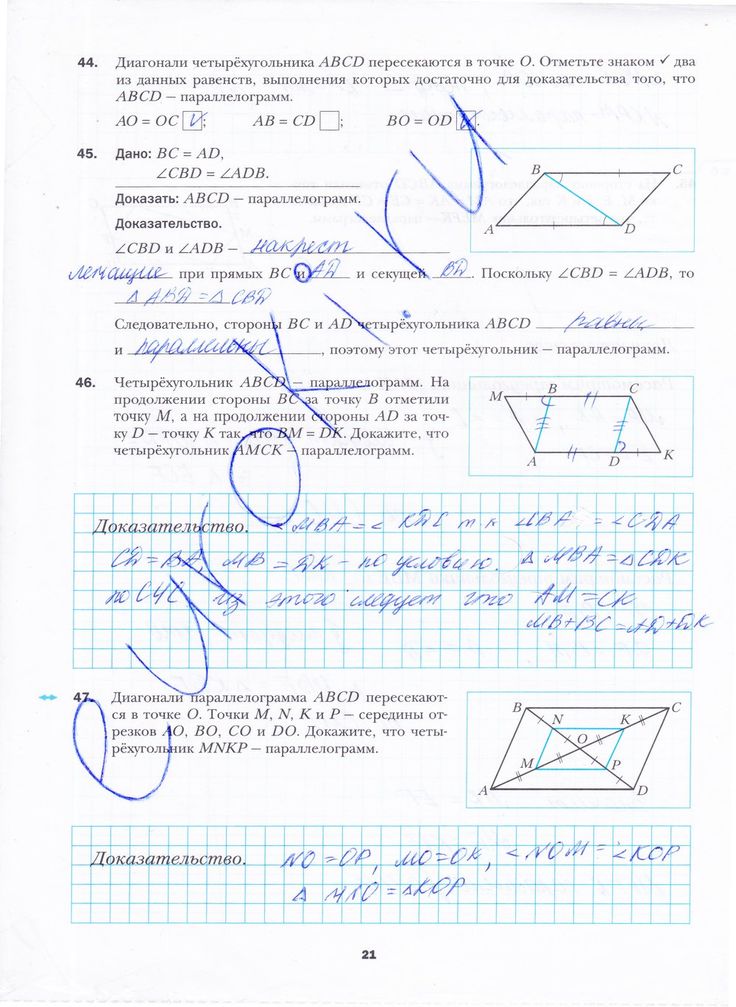 в период с 2016 по 2017 год. 14 (рис. 3). Участки исследования были выбраны так, чтобы отражать типичные сосновые/смешанные хвойные леса западной части Сьерра-Невады, в которых преобладала сосна пондероза 9.0145 14 . Участки были размещены в пораженных WPB сосновых/смешанных хвойных лесах на территории национальных лесов Эльдорадо, Станислав, Сьерра и Секвойя и были стратифицированы по высоте (914–1219 м, 1219–1524 м, 1524–1829 м над уровнем моря). ). В Национальном лесу Секвойя, самом южном национальном лесу в нашем исследовании, участки были стратифицированы с самой низкой полосой высот 1219–1524 м и расширены до верхней полосы высот 1829–2134 м, чтобы зафиксировать более похожий состав лесного сообщества как в более северных национальных лесах. Участки имеют различную структуру леса, а участки были выбраны в районах с базальной площадью сосны желтой > 35% и смертностью сосны желтой > 10%. На каждом участке вдоль трансект было заложено по пять круглых площадок площадью 0,041 га с расстоянием между площадками 80–200 м.
в период с 2016 по 2017 год. 14 (рис. 3). Участки исследования были выбраны так, чтобы отражать типичные сосновые/смешанные хвойные леса западной части Сьерра-Невады, в которых преобладала сосна пондероза 9.0145 14 . Участки были размещены в пораженных WPB сосновых/смешанных хвойных лесах на территории национальных лесов Эльдорадо, Станислав, Сьерра и Секвойя и были стратифицированы по высоте (914–1219 м, 1219–1524 м, 1524–1829 м над уровнем моря). ). В Национальном лесу Секвойя, самом южном национальном лесу в нашем исследовании, участки были стратифицированы с самой низкой полосой высот 1219–1524 м и расширены до верхней полосы высот 1829–2134 м, чтобы зафиксировать более похожий состав лесного сообщества как в более северных национальных лесах. Участки имеют различную структуру леса, а участки были выбраны в районах с базальной площадью сосны желтой > 35% и смертностью сосны желтой > 10%. На каждом участке вдоль трансект было заложено по пять круглых площадок площадью 0,041 га с расстоянием между площадками 80–200 м. В полевых условиях Феттиг и соавт. 14 нанес на карту все местоположения ствола относительно центра каждого участка, используя измерения азимута/расстояния. Принадлежность дерева к виду, высоте дерева и диаметру на высоте груди (DBH) регистрировали, если DBH превышал 6,35 см. Год гибели оценивали по цвету хвои и сохранению, если это произошло до закладки делянки и после этого непосредственно наблюдалось во время ежегодных посещений участка. Небольшой участок коры (~ 625 см 2 ) как с северной, так и с южной сторон был удален с мертвых деревьев, чтобы определить наличие ходов короедов. Форма, расположение и ориентация ходов обычно используются для различения видов короеда 18 . В некоторых случаях умершие жуки-короеды присутствовали под корой, чтобы дополнить идентификацию, основанную на формировании галереи. Весной и в начале лета 2018 года все полевые участки были повторно осмотрены, чтобы оценить, не упали ли мертвые деревья 14 .
В полевых условиях Феттиг и соавт. 14 нанес на карту все местоположения ствола относительно центра каждого участка, используя измерения азимута/расстояния. Принадлежность дерева к виду, высоте дерева и диаметру на высоте груди (DBH) регистрировали, если DBH превышал 6,35 см. Год гибели оценивали по цвету хвои и сохранению, если это произошло до закладки делянки и после этого непосредственно наблюдалось во время ежегодных посещений участка. Небольшой участок коры (~ 625 см 2 ) как с северной, так и с южной сторон был удален с мертвых деревьев, чтобы определить наличие ходов короедов. Форма, расположение и ориентация ходов обычно используются для различения видов короеда 18 . В некоторых случаях умершие жуки-короеды присутствовали под корой, чтобы дополнить идентификацию, основанную на формировании галереи. Весной и в начале лета 2018 года все полевые участки были повторно осмотрены, чтобы оценить, не упали ли мертвые деревья 14 .
 3: Сеть полевых участков охватывает 350-километровый градиент широты от Национального леса Эльдорадо на севере до Национального леса Секвойя на юге.
3: Сеть полевых участков охватывает 350-километровый градиент широты от Национального леса Эльдорадо на севере до Национального леса Секвойя на юге. Участки были стратифицированы по трем диапазонам высот в каждом лесу, при этом участки в Национальном лесу Секвойя (самый южный национальный лес) занимали диапазоны высот на 305 м выше трех диапазонов в других национальных лесах, чтобы охватить аналогичное сообщество сочинение.
Изображение полного размера
В типичном жизненном цикле WPB самки инициируют колонизацию хозяина, прокладывая туннели через внешнюю кору во флоэму и внешнюю ксилему, где они разрывают смоляные каналы. В результате живица выделяется и собирается на поверхности коры, что обычно наблюдается у других видов короедов. На ранних стадиях атаки самки выделяют компонент агрегационного феромона, который в сочетании с монотерпенами хозяина, высвобождаемыми из смоляных трубочек, привлекателен для сородичей 76 . Компонент антиагрегационного феромона вырабатывается на последних стадиях колонизации хозяина несколькими путями и, как полагают, снижает внутривидовую конкуренцию, изменяя поведение взрослых особей, чтобы свести к минимуму скученность развивающегося расплода внутри хозяина 77 . Было продемонстрировано, что летучие вещества из нескольких нехозяев, симпатричных с сосной пондероза, ингибируют привлечение WPB к его агрегационным феромонам 56,78 . В Калифорнии WPB обычно дает 2–3 поколения в течение одного года и часто может вытеснить других первичных жуков-короедов, таких как жук горной сосны, в соснах пондероза, особенно на более крупных деревьях 33 . Тем не менее темпы роста популяции WPB могут быть снижены из-за конкуренции с другими видами жуков, совместно обитающих на одном дереве-хозяине, а также из-за хищничества во время расселения в поисках хозяина 33 .
Было продемонстрировано, что летучие вещества из нескольких нехозяев, симпатричных с сосной пондероза, ингибируют привлечение WPB к его агрегационным феромонам 56,78 . В Калифорнии WPB обычно дает 2–3 поколения в течение одного года и часто может вытеснить других первичных жуков-короедов, таких как жук горной сосны, в соснах пондероза, особенно на более крупных деревьях 33 . Тем не менее темпы роста популяции WPB могут быть снижены из-за конкуренции с другими видами жуков, совместно обитающих на одном дереве-хозяине, а также из-за хищничества во время расселения в поисках хозяина 33 .
Сбор и обработка аэрофотоданных
Снимки в направлении надира были получены с помощью широкополосной красно-зелено-синей (RGB) камеры DJI Zenmuse X3 со стабилизацией на карданном подвесе 79 и стационарной мультиспектральной камеры Micasense Rededge3 с пятью узкими ленты 80 на самолете DJI Matrice 100 81 . Снимки были получены с обеих камер вдоль заранее запрограммированных аэрофотоснимков площадью около 40 га, окружающих каждый из 32 участков (каждый из которых содержит пять полевых участков), и были обработаны в несколько этапов, чтобы получить данные о структуре и составе местного леса, подходящие для наших статистических данных. анализы. Все изображения были сделаны в 2018 году в течение 3-месячного периода с начала апреля по начало июля, поэтому наша работа представляет собой вскрытие причин совокупной гибели деревьев. По призыву Wyngaard et al. 82 , мы устанавливаем «уровни продуктов данных», чтобы отразить конвейер обработки изображений от необработанных изображений (уровень 0) до калиброванной мелкомасштабной информации о структуре и составе леса на регулярной сетке (уровень 4), при этом каждый новый уровень данных получается из уровней ниже его. Здесь мы описываем этапы конвейера обработки и калибровки, представленные на рис. 4, и включаем дополнительные сведения в дополнительные методы.
анализы. Все изображения были сделаны в 2018 году в течение 3-месячного периода с начала апреля по начало июля, поэтому наша работа представляет собой вскрытие причин совокупной гибели деревьев. По призыву Wyngaard et al. 82 , мы устанавливаем «уровни продуктов данных», чтобы отразить конвейер обработки изображений от необработанных изображений (уровень 0) до калиброванной мелкомасштабной информации о структуре и составе леса на регулярной сетке (уровень 4), при этом каждый новый уровень данных получается из уровней ниже его. Здесь мы описываем этапы конвейера обработки и калибровки, представленные на рис. 4, и включаем дополнительные сведения в дополнительные методы.
Уровень 0 представляет необработанные данные с датчиков. Слева направо: фотография RGB с DJI Zenmuse X3, выходные изображения с Micasense Rededge3 (синий, зеленый, красный, ближний инфракрасный, красный край). Уровень 1 представляет основные результаты рабочего процесса SfM. Слева направо: плотное облако точек, ортофотоплан RGB, цифровая модель поверхности (DSM; высота земли плюс высота растительности). Уровень 2 представляет радиометрически или геометрически скорректированные продукты уровня 1. Слева направо: радиометрически скорректированная «красная» карта отражательной способности поверхности, радиометрически скорректированная «ближняя инфракрасная» карта отражательной способности поверхности, цифровая модель местности (ЦММ), полученная с помощью геометрической коррекции плотного облака точек, модель высоты полога (CHM; ЦММ вычитается из DSM). Уровень 3 представляет собой извлечение информации, относящейся к предметной области, из продуктов уровня 2 и делится на два подуровня. Продукты уровня 3а получаются с использованием только спектральных или только геометрических данных. Слева направо: карта нормализованного индекса различий растительности (NDVI) 83 , карта обнаруженных деревьев, полученная из CHM, обнаруженные деревья внутри красного многоугольника, многоугольники, представляющие сегментированные кроны деревьев, внутри красного многоугольника.
Уровень 1 представляет основные результаты рабочего процесса SfM. Слева направо: плотное облако точек, ортофотоплан RGB, цифровая модель поверхности (DSM; высота земли плюс высота растительности). Уровень 2 представляет радиометрически или геометрически скорректированные продукты уровня 1. Слева направо: радиометрически скорректированная «красная» карта отражательной способности поверхности, радиометрически скорректированная «ближняя инфракрасная» карта отражательной способности поверхности, цифровая модель местности (ЦММ), полученная с помощью геометрической коррекции плотного облака точек, модель высоты полога (CHM; ЦММ вычитается из DSM). Уровень 3 представляет собой извлечение информации, относящейся к предметной области, из продуктов уровня 2 и делится на два подуровня. Продукты уровня 3а получаются с использованием только спектральных или только геометрических данных. Слева направо: карта нормализованного индекса различий растительности (NDVI) 83 , карта обнаруженных деревьев, полученная из CHM, обнаруженные деревья внутри красного многоугольника, многоугольники, представляющие сегментированные кроны деревьев, внутри красного многоугольника. Продукты уровня 3b получаются с использованием как спектральных, так и геометрических данных. Слева направо: деревья, классифицированные как живые или мертвые на основе спектральной отражательной способности в каждой сегментированной кроне дерева, деревья, классифицированные как принимающие/не принимающие WPB. Уровень 4 представляет собой агрегирование продуктов уровня 3 в регулярные сетки, которые лучше отражают размер зерна проверки (например, для соответствия площади участков поля проверки) или которые предоставляют информацию в масштабе окрестности, а не в индивидуальном масштабе (например, доля на уровне насаждения). деревьев-хозяев). Слева направо: сетка, представляющая долю мертвых деревьев на ячейку, сетка, представляющая долю хостов на ячейку, сетка, представляющая среднюю высоту хосты на ячейку, плотность деревьев на ячейку. Все камеры имеют размер 20 × 20 м.
Продукты уровня 3b получаются с использованием как спектральных, так и геометрических данных. Слева направо: деревья, классифицированные как живые или мертвые на основе спектральной отражательной способности в каждой сегментированной кроне дерева, деревья, классифицированные как принимающие/не принимающие WPB. Уровень 4 представляет собой агрегирование продуктов уровня 3 в регулярные сетки, которые лучше отражают размер зерна проверки (например, для соответствия площади участков поля проверки) или которые предоставляют информацию в масштабе окрестности, а не в индивидуальном масштабе (например, доля на уровне насаждения). деревьев-хозяев). Слева направо: сетка, представляющая долю мертвых деревьев на ячейку, сетка, представляющая долю хостов на ячейку, сетка, представляющая среднюю высоту хосты на ячейку, плотность деревьев на ячейку. Все камеры имеют размер 20 × 20 м.
Полноразмерное изображение
Уровень 0: Необработанные данные с датчиков
Необработанные данные включали ~1900 изображений на объектив камеры (один широкополосный RGB-объектив и пять узкополосных мультиспектральных линз) для каждого из 32 участков (рис. 4). ; Уровень 0; Дополнительные рисунки 4 и 5). Перед аэрофотосъемкой две полоски ярко-оранжевой ткани (~ 100 × 15 см) были расположены в виде буквы «X» над постоянными памятниками, отмечая центры 5 полевых участков из Fettig et al. 14 (дополнительный рис. 6).
4). ; Уровень 0; Дополнительные рисунки 4 и 5). Перед аэрофотосъемкой две полоски ярко-оранжевой ткани (~ 100 × 15 см) были расположены в виде буквы «X» над постоянными памятниками, отмечая центры 5 полевых участков из Fettig et al. 14 (дополнительный рис. 6).
Мы предварительно запрограммировали воздушные разрезы с севера на юг с помощью Map Pilot для DJI в программном обеспечении iOS 84 на высоте 120 м над уровнем земли (с «землей», определенной с помощью цифровой модели рельефа с точностью до 1 угловой секунды 85 ) . Результирующее расстояние выборки земли составило ~5 см/пиксель для камеры Zenmuse X3 RGB и ~8 см/пиксель для мультиспектральной камеры Rededge3. Мы использовали 91,6% перекрытия изображений (вперед и сбоку) на земле для камеры Zenmuse X3 RGB и 83,9% перекрытия (вперед и сбоку) для мультиспектральной камеры Rededge3.
Уровень 1: основные результаты фотограмметрической обработки
Мы использовали фотограмметрию SfM, реализованную в Pix4Dmapper Cloud (www. pix4d.com), для создания плотных облаков точек (рис. 4; уровень 1, слева; дополнительный рис. 7), ортофотоплан ( Рис. 4; уровень 1, центральный и дополнительный рис. 8) и цифровые модели поверхности (рис. 4; уровень 1, правый и дополнительный рис. 9) для каждого полевого участка 71 . Для 29 сайтов мы обработали только мультиспектральные изображения Rededge3, чтобы создать эти продукты. Для трех участков мы обработали RGB и мультиспектральные изображения вместе, чтобы увеличить плотность точек плотного облака точек. Все проекты SfM привели к единому «блоку» обработки, что указывает на то, что все изображения в проекте были оптимизированы и обработаны вместе. Плотное облако точек представляет 9Координаты 0604 x , y и z , а также цвет миллионов точек на сайте. Ортомозаика представляет собой радиометрически некалиброванный вид сверху вниз на участок съемки, который сохраняет относительные положения объектов в пространстве 90 604 x 90 605 – 90 604 y 90 605. Цифровая модель поверхности представляет собой растровую версию плотного облака точек, которая показывает высоту над уровнем моря для каждого пикселя в сцене на расстоянии выборки земли от камеры, сгенерировавшей данные уровня 0.
pix4d.com), для создания плотных облаков точек (рис. 4; уровень 1, слева; дополнительный рис. 7), ортофотоплан ( Рис. 4; уровень 1, центральный и дополнительный рис. 8) и цифровые модели поверхности (рис. 4; уровень 1, правый и дополнительный рис. 9) для каждого полевого участка 71 . Для 29 сайтов мы обработали только мультиспектральные изображения Rededge3, чтобы создать эти продукты. Для трех участков мы обработали RGB и мультиспектральные изображения вместе, чтобы увеличить плотность точек плотного облака точек. Все проекты SfM привели к единому «блоку» обработки, что указывает на то, что все изображения в проекте были оптимизированы и обработаны вместе. Плотное облако точек представляет 9Координаты 0604 x , y и z , а также цвет миллионов точек на сайте. Ортомозаика представляет собой радиометрически некалиброванный вид сверху вниз на участок съемки, который сохраняет относительные положения объектов в пространстве 90 604 x 90 605 – 90 604 y 90 605. Цифровая модель поверхности представляет собой растровую версию плотного облака точек, которая показывает высоту над уровнем моря для каждого пикселя в сцене на расстоянии выборки земли от камеры, сгенерировавшей данные уровня 0.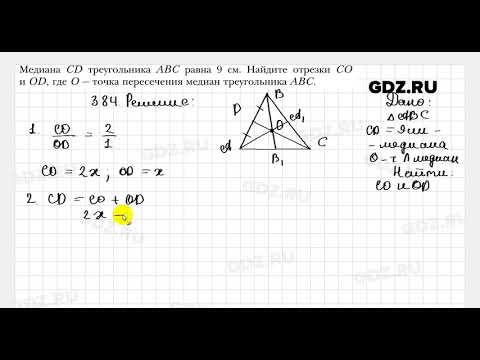
Уровень 2: скорректированные результаты фотограмметрической обработки
Радиометрические поправки
Карта коэффициента отражения с радиометрической поправкой (рис. 4; уровень 2, два рисунка слева; т. е. скорректированная версия ортофотоплана уровня 1; дополнительный рис. 10) была сгенерировано с помощью программного обеспечения Pix4D путем включения условий поступающего света для каждой узкой полосы камеры Rededge3 (захваченных одновременно с камерой Rededge3 с использованием встроенного датчика нисходящего света), а также предполетного изображения калибровочной панели с известным коэффициентом отражения (см. Дополнительный Таблица 3 для подробной информации о камере и панели калибровки).
Геометрические поправки
Мы внесли геометрическую поправку в плотное облако точек уровня 1 и цифровую модель поверхности, нормализовав эти данные для местности под растительностью. Мы создали цифровую модель местности, представляющую землю под растительностью с разрешением 1 м (рис. 4; уровень 2, третье изображение и дополнительный рисунок 11), классифицировав плотное облако точек каждой области съемки на «наземные» и «неназемные». ” точек с использованием алгоритма фильтра имитации ткани 86 реализовано в пакете lidR 53 и растрирование наземных точек с использованием растрового пакета 87 . Мы создали модель высоты навеса (рис. 4; уровень 2, четвертое изображение и дополнительный рис. 12), вычитая цифровую модель местности из цифровой модели поверхности.
4; уровень 2, третье изображение и дополнительный рисунок 11), классифицировав плотное облако точек каждой области съемки на «наземные» и «неназемные». ” точек с использованием алгоритма фильтра имитации ткани 86 реализовано в пакете lidR 53 и растрирование наземных точек с использованием растрового пакета 87 . Мы создали модель высоты навеса (рис. 4; уровень 2, четвертое изображение и дополнительный рис. 12), вычитая цифровую модель местности из цифровой модели поверхности.
Уровень 3: Извлечение информации, относящейся к предметной области
Уровень 3a: Данные, полученные из спектрального или геометрического продукта Уровня 2
Используя только спектральную информацию из радиометрически скорректированных карт отражения, мы рассчитали несколько вегетационных индексов, включая нормализованный разностный вегетативный индекс 83 (NDVI; рис. 4; уровень 3а, первое изображение и дополнительный рис. 13), нормализованная разность, красный край 88 (NDRE), красно-зеленый индекс 89 (RGI), красный край, хлорофилл индекс 90 (ДИ красный край ), и зеленый индекс хлорофилла 90 (ДИ зеленый ).
Используя только геометрическую информацию из модели высоты полога или плотного облака точек, нормализованного к рельефу, мы создали карты обнаруженных деревьев (рис. 4; уровень 3a, второе и третье изображения и дополнительный рисунок 14), протестировав в общей сложности 7 алгоритмы автоматического обнаружения деревьев и всего 177 наборов параметров (таблица 2). Мы использовали данные полевого участка для оценки каждого набора алгоритмов/параметров обнаружения деревьев путем преобразования измерений расстояния от центра и азимута деревьев на полевых участках в x – y позиций относительно центров полевых графиков, различимых на картах отражательной способности уровня 2 в виде оранжевых крестиков ткани, которые мы раскладывали перед каждым полетом. На картах отражения мы обнаружили 110 из 160 полевых центров делянок, в то время как некоторые центры делянок были скрыты из-за плотных переплетений крон деревьев или потому, что центр делянок находился непосредственно под кроной одного дерева. Для каждой из 110 полевых площадок с идентифицируемыми центрами делянок — «проверочные полевые делянки» — мы рассчитали 7 показателей структуры леса, используя наземные данные, собранные Fettig et al. 14 : общее количество деревьев, количество деревьев высотой более 15 м, средняя высота деревьев, высота дерева 25-го процентиля, высота дерева 75-го процентиля, среднее расстояние до ближайшего соседнего дерева и среднее расстояние до второго ближайшего соседа. Для каждого алгоритма обнаружения деревьев и набора параметров, описанных выше, мы рассчитали один и тот же набор из 7 структурных метрик в пределах охвата участков поля проверки. Мы рассчитали корреляцию Пирсона и среднеквадратичную ошибку (RMSE) между наземными данными и аэрофотоснимками для каждой из 7 структурных метрик для каждого из 177 алгоритмов/наборов параметров автоматического обнаружения деревьев. Для каждого алгоритма и набора параметров мы рассчитали его производительность по сравнению с другими алгоритмами в зависимости от того, находилась ли его корреляция Пирсона в пределах 5% от самой высокой корреляции Пирсона, а также находилась ли его среднеквадратическая ошибка в пределах 5% от самой низкой среднеквадратичной ошибки.
Для каждой из 110 полевых площадок с идентифицируемыми центрами делянок — «проверочные полевые делянки» — мы рассчитали 7 показателей структуры леса, используя наземные данные, собранные Fettig et al. 14 : общее количество деревьев, количество деревьев высотой более 15 м, средняя высота деревьев, высота дерева 25-го процентиля, высота дерева 75-го процентиля, среднее расстояние до ближайшего соседнего дерева и среднее расстояние до второго ближайшего соседа. Для каждого алгоритма обнаружения деревьев и набора параметров, описанных выше, мы рассчитали один и тот же набор из 7 структурных метрик в пределах охвата участков поля проверки. Мы рассчитали корреляцию Пирсона и среднеквадратичную ошибку (RMSE) между наземными данными и аэрофотоснимками для каждой из 7 структурных метрик для каждого из 177 алгоритмов/наборов параметров автоматического обнаружения деревьев. Для каждого алгоритма и набора параметров мы рассчитали его производительность по сравнению с другими алгоритмами в зависимости от того, находилась ли его корреляция Пирсона в пределах 5% от самой высокой корреляции Пирсона, а также находилась ли его среднеквадратическая ошибка в пределах 5% от самой низкой среднеквадратичной ошибки. Мы суммировали количество метрик структуры леса, для которых он достиг этих 5% пороговых значений для каждого алгоритма/набора параметров. Для автоматического обнаружения деревьев по всему исследованию мы выбрали набор алгоритмов/параметров, который показал хорошие результаты по большинству показателей леса (см. раздел «Результаты»).
Мы суммировали количество метрик структуры леса, для которых он достиг этих 5% пороговых значений для каждого алгоритма/набора параметров. Для автоматического обнаружения деревьев по всему исследованию мы выбрали набор алгоритмов/параметров, который показал хорошие результаты по большинству показателей леса (см. раздел «Результаты»).
Полноразмерная таблица
Мы очертили кроны отдельных деревьев (рис. 4; уровень 3a, четвертое изображение и дополнительный рис. 15) с помощью алгоритма сегментации водоразделов, контролируемого маркерами, 99 , реализованного в пакете ForestTools 97 , с использованием обнаруженных верхушек деревьев. как маркеры. Если алгоритму автоматической сегментации не удавалось сгенерировать сегмент кроны для обнаруженного дерева (например, часто коряги с очень маленьким отпечатком кроны), генерировалась круглая крона с радиусом 0,5 м. Если сегментация генерировала несколько полигонов для одного обнаруженного дерева, сохранялся только полигон, содержащий обнаруженное дерево. Поскольку перекрытие изображений уменьшается ближе к краям общей траектории полета и снижает качество обработки ССМ в этих областях, мы исключили сегментированные кроны в пределах 35 м от края области съемки. Учитывая более узкое поле зрения мультиспектральной камеры Rededge3 по сравнению с RGB-камерой X3, оптические параметры которой использовались для определения области съемки ~40 га вокруг каждой площадки, а также дополнительную буферизацию в 35 м, площадь съемки на каждой площадке составляла ~ 30 га (дополнительная таблица 1).
Если сегментация генерировала несколько полигонов для одного обнаруженного дерева, сохранялся только полигон, содержащий обнаруженное дерево. Поскольку перекрытие изображений уменьшается ближе к краям общей траектории полета и снижает качество обработки ССМ в этих областях, мы исключили сегментированные кроны в пределах 35 м от края области съемки. Учитывая более узкое поле зрения мультиспектральной камеры Rededge3 по сравнению с RGB-камерой X3, оптические параметры которой использовались для определения области съемки ~40 га вокруг каждой площадки, а также дополнительную буферизацию в 35 м, площадь съемки на каждой площадке составляла ~ 30 га (дополнительная таблица 1).
Уровень 3b: Данные, полученные на основе спектральной и геометрической информации
Мы наложили сегментированные короны на карты отражательной способности с 20 участков, охватывающих широтный и высотный градиенты в исследовании. Используя QGIS (https://qgis.org/en/site/), мы вручную классифицировали 564 дерева как живые/мертвые и как один из 5 доминирующих видов в исследуемой области (сосна желтоватая, Pinus lambertiana , Abies concolor ). , Calocedrus decurrens или Quercus kelloggi ), используя в качестве ориентира нанесенные на карту наземные данные. Каждое дерево далее классифицировалось как «хозяин» для сосны пондероза или «нехозяин» для всех других видов9.0145 18 . Мы извлекли все значения пикселей внутри каждого сегментированного полигона кроны из пяти ортотрансформированных карт отражения уровня 2 (по одной на узкую полосу на камере Rededge3), а также из пяти карт индекса растительности уровня 3a с использованием пакета velox 100 . Для каждого многоугольника кроны мы рассчитали среднее значение извлеченных пикселей уровня 2 и уровня 3a и использовали их в качестве десяти независимых переменных в модели усиленной логистической регрессии с пятикратной перекрестной проверкой, чтобы предсказать, были ли деревья, классифицированные вручную, живыми или мертвыми. Только для живых деревьев мы аналогичным образом использовали все 10 средних значений коэффициента отражения на полигон кроны, чтобы предсказать виды деревьев с помощью пятикратного перекрестно проверенного регуляризованного дискриминантного анализа.
, Calocedrus decurrens или Quercus kelloggi ), используя в качестве ориентира нанесенные на карту наземные данные. Каждое дерево далее классифицировалось как «хозяин» для сосны пондероза или «нехозяин» для всех других видов9.0145 18 . Мы извлекли все значения пикселей внутри каждого сегментированного полигона кроны из пяти ортотрансформированных карт отражения уровня 2 (по одной на узкую полосу на камере Rededge3), а также из пяти карт индекса растительности уровня 3a с использованием пакета velox 100 . Для каждого многоугольника кроны мы рассчитали среднее значение извлеченных пикселей уровня 2 и уровня 3a и использовали их в качестве десяти независимых переменных в модели усиленной логистической регрессии с пятикратной перекрестной проверкой, чтобы предсказать, были ли деревья, классифицированные вручную, живыми или мертвыми. Только для живых деревьев мы аналогичным образом использовали все 10 средних значений коэффициента отражения на полигон кроны, чтобы предсказать виды деревьев с помощью пятикратного перекрестно проверенного регуляризованного дискриминантного анализа. Усиленная логистическая регрессия и регуляризованный дискриминантный анализ были реализованы с использованием пакета Caret в R 9.0145 101 . Мы использовали эти модели для классификации всех крон деревьев в наборе данных как живых или мертвых (рис. 4; уровень 3b, первое изображение и дополнительный рис. 16), а также для классификации видов живых деревьев (а затем хозяев или не- хост; рис. 4; уровень 3b, второе изображение; дополнительный рисунок 17).
Усиленная логистическая регрессия и регуляризованный дискриминантный анализ были реализованы с использованием пакета Caret в R 9.0145 101 . Мы использовали эти модели для классификации всех крон деревьев в наборе данных как живых или мертвых (рис. 4; уровень 3b, первое изображение и дополнительный рис. 16), а также для классификации видов живых деревьев (а затем хозяев или не- хост; рис. 4; уровень 3b, второе изображение; дополнительный рисунок 17).
Поскольку вершины мертвых деревьев без иголок узкие, они могут быть недостаточно хорошо представлены в облаках точек, полученных с помощью фотограмметрии SfM, что приводит к занижению оценок их высоты. Кроме того, полевые измерения могут завышать высоту живых деревьев по сравнению с методами аэрофотосъемки 9.0145 102 . Чтобы исправить эти погрешности измерений, мы откалибровали измерения высоты деревьев с воздуха по наземным измерениям высоты. В частности, мы определили кроны 451 дерева, измеренного в полевых условиях, в данных о деревьях, полученных с помощью дронов, смоделировали взаимосвязь между высотой деревьев, измеренных в полевых условиях и с помощью дронов, как для живых, так и для мертвых деревьев, и использовали модели для корректировки деревьев, измеренных с помощью дронов. высоты (дополнительные методы). Мы применили консервативную поправку на высоту к живым и мертвым деревьям, основываясь на том, что деревья, измеренные дроном, имели высоту более 20 м, что увеличило высоту мертвых деревьев в среднем на 2,8 м и уменьшило высоту живых деревьев в среднем на 0,9 м.м (дополнительные рисунки 18–20 и дополнительное примечание 2). Наконец, мы оценили базальную площадь каждого дерева по их скорректированной высоте, измеренной дроном, с использованием простой линейной регрессии для конкретных видов взаимосвязи между высотой и DBH, измеренной на совпадающих полевых графиках из Fettig et al. 14 .
высоты (дополнительные методы). Мы применили консервативную поправку на высоту к живым и мертвым деревьям, основываясь на том, что деревья, измеренные дроном, имели высоту более 20 м, что увеличило высоту мертвых деревьев в среднем на 2,8 м и уменьшило высоту живых деревьев в среднем на 0,9 м.м (дополнительные рисунки 18–20 и дополнительное примечание 2). Наконец, мы оценили базальную площадь каждого дерева по их скорректированной высоте, измеренной дроном, с использованием простой линейной регрессии для конкретных видов взаимосвязи между высотой и DBH, измеренной на совпадающих полевых графиках из Fettig et al. 14 .
Мы отмечаем, что наше исследование основано на создании продуктов уровня 3a для их объединения и создания продуктов уровня 3b, таких как классифицированные древовидные карты, но это не обязательно так. Например, методы глубокого обучения/нейронных сетей могут одновременно использовать как спектральную, так и геометрическую информацию из продуктов данных более низкого уровня, чтобы находить и классифицировать деревья в сцене и напрямую генерировать продукты уровня 3b без необходимости сначала генерировать продукты уровня 3a. продукты 103 104 .
продукты 103 104 .
Уровень 4: Агрегирование в регулярные сетки
Мы растеризовали данные о структуре и составе леса с пространственным разрешением, аналогичным разрешению полевых графиков, чтобы лучше соответствовать размеру зерна, при котором мы проверяли алгоритмы автоматического обнаружения деревьев. В каждой ячейке растра подсчитывались: количество сухих деревьев, количество сосны желтой, общее количество деревьев и средняя высота сосен желтых. Значения этих переменных в каждой ячейке сетки и производные от них использовались для визуализации и моделирования. Здесь мы показываем долю мертвых деревьев на ячейку (рис. 4; уровень 4, первое изображение и дополнительный рисунок 21), долю деревьев-хозяев на ячейку (рис. 4; уровень 4, второе изображение), среднюю высоту деревьев пондероза в каждой ячейке (рис. 4; уровень 4, третье изображение) и общее количество деревьев на ячейку (рис. 4; уровень 4, четвертое изображение).
Примечание о предположениях о мертвых деревьях
Для целей данного исследования мы предположили, что все мертвые деревья были сосной пондероза и, таким образом, хозяевами, колонизированными WPB.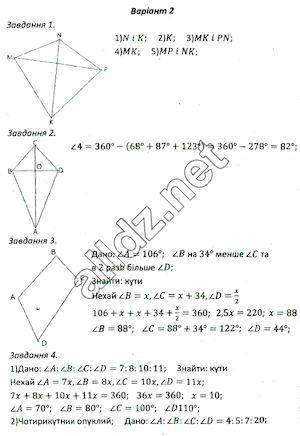 Это достаточно хорошее предположение для области нашего исследования; например, Fettig et al. 14 обнаружили, что 73,4% мертвых деревьев на совпадающих участках поля были сосной желтой. Смертность была сосредоточена в классах с большим диаметром и в основном относилась к WPB (см. рис. 5 Fettig et al. 14 ). Следующей по величине долей сухих деревьев был кедр ладан, на долю которого приходилось 18,72% сухих деревьев на полевых участках. Хотя обнаруженная смертность, скорее всего, связана с уничтожением сосны пондерозы WPB, очень важно интерпретировать наши результаты с учетом этих ограничений.
Это достаточно хорошее предположение для области нашего исследования; например, Fettig et al. 14 обнаружили, что 73,4% мертвых деревьев на совпадающих участках поля были сосной желтой. Смертность была сосредоточена в классах с большим диаметром и в основном относилась к WPB (см. рис. 5 Fettig et al. 14 ). Следующей по величине долей сухих деревьев был кедр ладан, на долю которого приходилось 18,72% сухих деревьев на полевых участках. Хотя обнаруженная смертность, скорее всего, связана с уничтожением сосны пондерозы WPB, очень важно интерпретировать наши результаты с учетом этих ограничений.
Данные об окружающей среде
Мы использовали CWD 105 из среднего значения модели характеристики бассейна 106 за 1981–2010 гг. в качестве интегральной меры исторических условий температуры и влажности для каждого из 32 участков. Более высокие значения CWD соответствуют исторически более жарким и сухим условиям, а более низкие значения соответствуют исторически более холодным и влажным условиям. Было показано, что CWD хорошо коррелирует с широким спектром гибели деревьев в Сьерра-Неваде 11 , а также гибель деревьев, вызванная короедом 107 . Леса вдоль всего градиента CWD, используемого в этом исследовании, испытали исключительную жаркую засуху в период с 2012 по 2016 год с серьезностью, по крайней мере, 1200-летнего события и, возможно, более серьезного, чем 10000-летнее событие 2,3 . Мы преобразовали значение CWD для каждого участка в 90 604 z 90 605 баллов, представляющих отклонение этого участка от среднего значения CWD по всему климатическому диапазону сосны Сьерра-Невада пондероза, как определено из 179гербарные записи, описанные в Baldwin et al. 108 . Таким образом, CWD z -балл, равный 1, будет означать, что CWD в этом месте на одно стандартное отклонение жарче/суше, чем среднее значение CWD по всем геолоцированным гербарным записям для сосны пондерозовой в Сьерра-Неваде.
Было показано, что CWD хорошо коррелирует с широким спектром гибели деревьев в Сьерра-Неваде 11 , а также гибель деревьев, вызванная короедом 107 . Леса вдоль всего градиента CWD, используемого в этом исследовании, испытали исключительную жаркую засуху в период с 2012 по 2016 год с серьезностью, по крайней мере, 1200-летнего события и, возможно, более серьезного, чем 10000-летнее событие 2,3 . Мы преобразовали значение CWD для каждого участка в 90 604 z 90 605 баллов, представляющих отклонение этого участка от среднего значения CWD по всему климатическому диапазону сосны Сьерра-Невада пондероза, как определено из 179гербарные записи, описанные в Baldwin et al. 108 . Таким образом, CWD z -балл, равный 1, будет означать, что CWD в этом месте на одно стандартное отклонение жарче/суше, чем среднее значение CWD по всем геолоцированным гербарным записям для сосны пондерозовой в Сьерра-Неваде.
Статистическая модель
Мы использовали обобщенную линейную модель с нулевым биномиальным откликом и логит-связью для прогнозирования вероятности гибели сосны желтовато-зеленой в каждой ячейке 20 × 20 м с использованием общего количества сосен пондерозовых в каждой ячейке как количество испытаний, а количество мертвых деревьев в каждой ячейке как количество «успехов». В качестве ковариант мы использовали долю деревьев-хозяев WPB (т. е. сосны пондерозы) в каждой ячейке, среднюю высоту деревьев сосны пондерозы в каждой ячейке, количество деревьев всех видов (общая плотность) в каждой ячейке и CWD на уровне сайта с использованием уравнения. 1. Обратите внимание, что двусторонняя взаимосвязь между общей плотностью и долей деревьев-хозяев прямо пропорциональна количеству пондерозовых сосен в ячейке. Мы центрировали и масштабировали все значения предикторов и использовали слабо регуляризирующие априорные значения по умолчанию из пакета brms 9.0145 109 . Чтобы измерить и учесть пространственную автокорреляцию, лежащую в основе смертности сосны пондерозной, мы разделили данные на каждом участке на случайную выборку из 200 ячеек размером 20 × 20 м, представляющих ~ 27,5% обследованной площади. Кроме того, с этими подвыборочными данными мы включили отдельный точный член гауссовского процесса для каждого участка нецентрированного/немасштабированного взаимодействия между x и y положением каждой ячейки, используя функцию gp() в пакете brms 109.
В качестве ковариант мы использовали долю деревьев-хозяев WPB (т. е. сосны пондерозы) в каждой ячейке, среднюю высоту деревьев сосны пондерозы в каждой ячейке, количество деревьев всех видов (общая плотность) в каждой ячейке и CWD на уровне сайта с использованием уравнения. 1. Обратите внимание, что двусторонняя взаимосвязь между общей плотностью и долей деревьев-хозяев прямо пропорциональна количеству пондерозовых сосен в ячейке. Мы центрировали и масштабировали все значения предикторов и использовали слабо регуляризирующие априорные значения по умолчанию из пакета brms 9.0145 109 . Чтобы измерить и учесть пространственную автокорреляцию, лежащую в основе смертности сосны пондерозной, мы разделили данные на каждом участке на случайную выборку из 200 ячеек размером 20 × 20 м, представляющих ~ 27,5% обследованной площади. Кроме того, с этими подвыборочными данными мы включили отдельный точный член гауссовского процесса для каждого участка нецентрированного/немасштабированного взаимодействия между x и y положением каждой ячейки, используя функцию gp() в пакете brms 109. . Гауссовский процесс оценивает пространственную ковариацию в переменной отклика (логарифмические шансы гибели сосны желтой) вместе с эффектами других ковариат.
. Гауссовский процесс оценивает пространственную ковариацию в переменной отклика (логарифмические шансы гибели сосны желтой) вместе с эффектами других ковариат.
$$\begin{array}{*{20}{l}} \hfill {y_{i,j} \sim} & \hfill {\left\{ {\begin{array}{*{20}{ l}} {0,} \hfill & p \hfill \\ {Binom(n_i,\pi _i),} \hfill & {1 — p} \hfill \end{массив}} \right.} & \hfill { \,}\\ \hfill {\ log it (\ pi _i) = } {\,} & \ hfill {\ beta _0 +} & \ hfill {\,} \\ \ hfill {\,} & \ hfill { \beta _1X_{cwd,j} + \beta _2X_{propHost,i} + \beta _3X_{PipoHeight,i} +} & \hfill {\,}\\ \hfill {\,} & \hfill {\beta _4X_ {overallDensity,i} + \beta _5X_{overallBA,i} +} & \hfill {\,}\\ \hfill {\,} & \hfill {\beta _6X_{cwd,j}X_{PipoHeight,i} + \beta _7X_{cwd,j}X_{propHost,i} +} & \hfill {\,}\\ \hfill {\,} & \hfill {\beta _8X_{cwd,j}X_{overallDensity,i} + \бета _9X_ {cwd, j} X_ {overallBA, i} +} & \ hfill {\,} \\ \ hfill {\,} & \ hfill {\ beta _ {10} X_ {propHost, i} X_ {PipoHeight, i } + \beta _{11}X_{propHost,i}X_{overallDensity,i} +} & \hfill {\,}\\ \hfill {\,} & \hfill {\beta _{12}X_{PipoHeight ,i}X_{overallBA,i} +} & \hfill {\,}\\ \hfill {\,} & \hfill {\beta _{13}X_{cwd,j}X_{propHost,i}X_{ PipoHeight,i} +} & \hfill {\,}\\ \hfill {\,} & \hfill {{\cal{G}}{\cal{P}}_j(x_i,y_i)} & \hfill { \,}\end{массив}$$
(1)
Где y i — количество мертвых деревьев в ячейке i , n i — сумма мертвых деревьев (предполагается, что это сосна пондероза) и живых деревьев в ячейке 4 сосны 60 i , \(\pi _i\) — вероятность гибели деревьев сосны пондерозы в ячейке i , p — вероятность того, что в ячейке, возникшей в результате независимого немоделируемого процесса, не будет мертвых деревьев, \(X_{cwd,j}\) — это z -оценка CWD для сайта j , \(X_{propHost,i}\) — масштабированная доля деревьев, представляющих собой сосну пондерозу, в ячейке i , \(X_{PipoHeight,i}\) — масштабированная средняя высота сосны пондероза в ячейке i , \(X_{overallDensity,i}\) — масштабированная плотность всех деревьев в ячейке i , \(X_{overallBA,i}\) — масштабированная базальная площадь всех деревьев в ячейке i , x i и y i — координаты x и y центра тяжести ячейки в системе отсчета координат EPSG3310, а GPj представляет собой точный гауссовский процесс, описывающий пространственную ковариацию между ячейками в точке j .
Мы подгоняем эту модель с помощью пакета brms 109 , который реализует расширение No U-Turn Sampler для гамильтонового алгоритма Монте-Карло 110 на языке программирования Stan 111 . Мы использовали 4 цепочки по 5000 итераций каждая (2000 прогревов, 3000 проб) и подтвердили сходимость цепей, убедившись, что все значения Rhat меньше 1,19.0145 112 и что объемные и хвостовые эффективные размеры выборки (ESS) для каждого оцениваемого параметра более чем в 100 раз превышали количество цепочек (т.е.> 400 в нашем случае). Мы использовали апостериорные прогностические проверки для визуального подтверждения эффективности модели путем наложения кривых плотности предсказанного количества мертвых деревьев на ячейку по наблюдаемому числу 113 . Для апостериорных прогностических проверок мы использовали 50 случайных выборок из модели, подходящей для создания 50 кривых плотности, и гарантировали, что кривые были центрированы по наблюдаемому распределению, уделяя особое внимание производительности модели при нулевом счете (дополнительный рисунок 22).
Доступность данных
Все полевые данные и данные с дронов, обработанные для этого исследования, доступны через Open Science Framework по адресу https://doi.org/10.17605/OSF.IO/3CWF9 114 . Файл административных границ Лесной службы Министерства сельского хозяйства США (S_USA.AdministrativeForest.shp) можно найти по адресу https://data.fs.usda.gov/geodata/edw/datasets.php?dsetCategory=boundaries. Версию 30-летних исторических данных о среднем климатическом дефиците воды за 1981–2010 годы (cwd1981_2010_ave_HST_1550861123.tif) за 2014 год можно найти на сайте California Climate Commons по адресу http://climate.calcommons.org/dataset/2014-CA-BCM. Набор данных, представляющих географические местоположения сосны пондерозы, полученные из гербарных записей (California_Species_clean_All_epsg_3310.csv), можно найти по адресу https://doi.org/10.6078/D16K5W 9.0145 115 . Векторный файл, представляющий географические подразделения Джепсона в Калифорнии и используемый для определения региона Сьерра-Невада, можно запросить по адресу https://ucjeps. berkeley.edu/eflora/geography.html.
berkeley.edu/eflora/geography.html.
Наличие кода
Статистический анализ выполнен с использованием пакетов brms. За исключением программного обеспечения SfM (Pix4Dmapper Cloud) и программного обеспечения ГИС QGIS, вся обработка данных и анализ были выполнены с использованием R 116 . Весь код, используемый для получения результатов этого исследования, доступен через GitHub по адресу и отражен в Open Science Framework по адресу https://doi.org/10.17605/OSF.IO/WPK5Z 9.0145 117 .
Ссылки
USDAFS. Пресс-релиз: Исследование показало, что в 2018 году в Калифорнии погибло 18 миллионов деревьев . https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/FSEPRD609321.pdf (USDAFS, 2019).
Гриффин Д. и Анчукайтис К. Дж. Насколько необычна засуха в Калифорнии в 2012–2014 гг.? Геофиз. Рез. лат. 41 , 9017–9023 (2014).
ОБЪЯВЛЕНИЕ Статья Google ученый
Asner, G. P. et al. Прогрессирующая потеря воды в пологе леса во время засухи в Калифорнии в 2012-2015 гг. Проц. Натл акад. науч. США 113 , E249–E255 (2016).
КАС пабмед Статья Google ученый
Бродрик, П. Г. и Аснер, Г. П. Предикторы гибели хвойных деревьев с помощью дистанционного зондирования во время сильной засухи. Окружающая среда. Рез. лат. 12 , 115013 (2017).
ОБЪЯВЛЕНИЕ Статья Google ученый
Феттиг, Си Джей в Управление лесами Сьерра-Невады. PSW-GTR-237 Гл. 2 (Лесная служба Министерства сельского хозяйства США, 2012 г.
).Колб, Т. Э. и др. Наблюдаемое и ожидаемое воздействие засухи на лесных насекомых и болезни в США. Для. Экол. Управление 380 , 321–334 (2016).
Артикул Google ученый
Waring, R. H. & Pitman, G. B. Модификация насаждений скальной сосны для изменения восприимчивости к нападению лубоеда горной сосны. Экология 66 , 889–897 (1985).
Артикул Google ученый
Restaino, C. et al. Структура леса и климат опосредуют вызванную засухой гибель деревьев в лесах Сьерра-Невады, США. Экол. заявл. 0 , e01902 (2019).
Артикул Google ученый
USDAFS. Пресс-релиз: Рекордные 129 миллионов мертвых деревьев в Калифорнии . https://www.fs.usda.gov/Internet/FSE_DOCUMENTS/fseprd566303.
pdf (USDAFS, 2017).Young, D. J. N. et al. Долговременный климат и конкуренция объясняют характер гибели лесов в условиях сильной засухи. Экол. лат. 20 , 78–86 (2017).
ПабМед Статья Google ученый
Raffa, K.F. et al. Межмасштабные факторы природных нарушений, склонных к антропогенному усилению: динамика извержений короедов. BioScience 58 , 501–517 (2008).
Артикул Google ученый
Boone, C.K., Aukema, B.H., Bohlmann, J., Carroll, A.L. & Raffa, K.F. Эффективность физиологии защиты деревьев зависит от плотности популяции жуков-короедов: основа для положительной обратной связи у эруптивных видов. Кан. Дж. Рез. 41 , 1174–1188 (2011).
Артикул Google ученый
Стефенсон, Н. Л., Дас, А. Дж., Амперзее, Н. Дж. и Булаон, Б. М. Какие деревья погибают во время засухи? Ключевая роль выбора насекомых-хозяев. Ж. Экол. 75 , 2383–2401 (2019).
Артикул Google ученый
Сенф К., Кэмпбелл Э. М., Пфлугмахер Д., Вулдер М. А. и Хостерт П. Многомасштабный анализ динамики вспышек западной еловой листовертки. Ландск. Экол. 32 , 501–514 (2017).
Артикул Google ученый
Seidl, R. et al. Мелкий жук, масштабные драйверы: как региональные и ландшафтные факторы влияют на вспышки европейского елового короеда.
J. Appl Ecol. 53 , 530–540 (2016).Артикул Google ученый
Fettig, CJ в Insects and Diseases of Mediterranean Forest Systems (ред. Lieutier, F. & Paine, TD) 499–528 (Springer International Publishing, 2016).
Раффа, К. Ф. и Берриман, А. А. Роль устойчивости растений-хозяев в колонизационном поведении и экологии жуков-короедов (Coleoptera: Scolytidae). Экол. моногр. 53 , 27–49 (1983).
Артикул Google ученый
Логан, Дж. А., Уайт, П., Бенц, Б. Дж. и Пауэлл, Дж. А. Модельный анализ пространственных закономерностей вспышек горного соснового жука. Теор. Народ. биол. 53 , 236–255 (1998).
КАС пабмед МАТЕМАТИКА Статья Google ученый
Франчески В. Р., Крокене П., Кристиансен Э. и Креклинг Т. Анатомическая и химическая защита коры хвойных деревьев от короедов и других вредителей. Н. Фитол. 167 , 353–376 (2005).
КАС Статья Google ученый
Раффа К.Ф., Грегуар Ж.-К. и Стаффан Линдгрен, Б. Естественная история и экология короедов 1–40 (Elsevier, 2015).
Bentz, B.J. et al. Изменение климата и короеды западной части США и Канады: прямое и косвенное воздействие. BioScience 60 , 602–613 (2010).
Артикул Google ученый
Харт, С. Дж., Веблен, Т. Т., Шнайдер, Д. и Молоч, Н. П. Летняя и зимняя засухи вызывают возникновение и распространение вспышки елового жука. Экология 98 , 2698–2707 (2017).
ПабМед Статья Google ученый
Нетерер, С., Панассити, Б., Пеннерсторфер, Дж. и Мэтьюз, Б. Острая засуха Является важным фактором заражения короедом ельников австрийской. Фронт. За. Глоб. Изменение 2 , 39 (2019).
Кайзер, К. Э., МакГлинн, Б. Л. и Эмануэль, Р. Э. Экогидрология очага: горный сосновый лубоед в первую очередь поражает деревья в более сухих местах ландшафта.
Экогидрология 6 , 444–454 (2013).Артикул Google ученый
Марини Л. и др. Климатические факторы динамики вспышек короеда в еловых лесах обыкновенной. Экография 40 , 1426–1435 (2017).
Артикул Google ученый
Sambaraju, K.R., Carroll, A.L. & Aukema, B.H. Многолетние погодные аномалии, связанные со сдвигами ареалов горного соснового лубоеда перед крупными эпидемиями. Для. Экол. Управление 438 , 86–95 (2019).
Артикул Google ученый
Hayes, C.J., Fettig, C.J. & Merrill, L.D. Оценка множественных воронкообразных ловушек и характеристик насаждений для оценки смертности деревьев, вызванной западным сосновым жуком. Ж. эконом. Энтомол. 102 , 2170–2182 (2009).
ПабМед Статья Google ученый
Миллер, Дж. М. и Кин, Ф. П. Биология западного соснового жука и борьба с ним: краткое изложение первых пятидесяти лет исследований (Министерство сельского хозяйства США, 1960 г.).
Чубатый А. М., Ройтберг Б. Д. и Ли К. Модель динамического выбора хозяев для горного соснового лубоеда, Dendroctonus ponderosae Hopkins. Экол. Модель. 220 , 1241–1250 (2009).
Артикул Google ученый
Граф М., Рейд М. Л., Аукема Б. Х. и Линдгрен Б. С. Связь диаметра дерева с размером тела и содержанием липидов у жуков горной сосны. Кан. Энтомол. 144 , 467–477 (2012).
Артикул Google ученый
Кляйн, У. Х., Паркер, Д. Л. и Дженсен, К. Э. Тенденции нападения, появления и истощения насаждений горного соснового лубоеда в древостоях скрученной сосны во время вспышки. Окружающая среда. Энтомол. 7 , 732–737 (1978).
Артикул Google ученый
Митчелл, Р. Г. и Прейслер, Х. К. Анализ пространственных закономерностей скальной сосны, атакованной вспышками популяций лубоеда горной сосны. Для. науч. 37 , 1390–1408 (1991).
Google ученый
Прейслер, Х.К. Моделирование пространственных структур деревьев, атакованных короедом. Заявл. Стат. 42 , 501 (1993).
МАТЕМАТИКА Статья Google ученый
Jactel, H. & Brockerhoff, E.G. Разнообразие деревьев снижает травоядность лесных насекомых. Экол. лат. 10 , 835–848 (2007).
ПабМед Статья Google ученый
Faccoli, M. & Bernardinelli, I. Состав и высота еловых лесов влияют на восприимчивость к нападениям короедов: последствия для управления лесами. Леса 5 , 88–102 (2014).
Артикул Google ученый
Берриман, А. А. в Короеды в хвойных деревьях Северной Америки: система изучения эволюционной биологии 264–314 (University of Texas Press, 1982).
Fettig, C.J. et al. Эффективность методов управления растительностью для предотвращения и борьбы с инвазиями короедов в хвойных лесах на западе и юге США.
Для. Экол. Управление 238 , 24–53 (2007).Артикул Google ученый
Моек, Х. А., Вуд, Д. Л. и Линдал, К. К. Поведение короедов (Coleoptera: Scolytidae) при выборе хозяина, атакующих Pinus ponderosa , с особым акцентом на западном сосновом жуке, Dendroctonus brevicomis . J. Chem. Экол. 7 , 49–83 (1981).
КАС пабмед Статья Google ученый
Эвенден, М. Л., Уайтхаус, К. М. и Сайкс, Дж. Факторы, влияющие на летную способность жука горной сосны (Coleoptera: Curculionidae: Scolytinae). Окружающая среда. Энтомол. 43 , 187–196 (2014).
КАС пабмед Статья Google ученый
Раффа, К. Ф. и Берриман, А. А. Накопление монотерпенов и связанных с ними летучих веществ после инокуляции пихты великой грибком, передаваемым пихтовым резчиком, Scolytus ventralis (Coleoptera: Scolytidae).
Кан. Энтомол. 114 , 797–810 (1982).КАС Статья Google ученый
Anderegg, W. R. L. et al. Смертность деревьев от засухи, насекомых и их взаимодействие в условиях изменяющегося климата. Н. Фитол. 208 , 674–683 (2015).
Артикул Google ученый
Кейн, В. Р. и др. Оценка воздействия пожаров на пространственную структуру леса с использованием данных Landsat и бортовых LiDAR в национальном парке Йосемити. Дистанционный датчик окружающей среды. 151 , 89–101 (2014).
ОБЪЯВЛЕНИЕ Статья Google ученый
Ларсон, А. Дж. и Черчилль, Д. Пространственные модели деревьев в лесах с частыми пожарами на западе Северной Америки, включая механизмы формирования моделей и последствия для разработки методов сокращения расхода топлива и восстановления.
Для. Экол. Управление 267 , 74–92 (2012).Артикул Google ученый
Morris, J. L. et al. Управление воздействием жуков-короедов на экосистемы и общество: приоритетные вопросы для мотивации будущих исследований. J. Appl. Экол. 54 , 750–760 (2017).
Артикул Google ученый
Шикломанов А.Н. и др. Расширение экспериментов по глобальным изменениям за счет интеграции методов дистанционного зондирования. Фронт. Экол. Окружающая среда . 17 , 215–224 (2019).
Jeronimo, S. M. A. et al. Структура и структура леса различаются в зависимости от климата и формы рельефа в горных ландшафтах горной Сьерра-Невады. Для. Экол. Управление 437 , 70–86 (2019).
Артикул Google ученый
McDowell, N. et al. Механизмы выживания и гибели растений при засухе: почему одни растения выживают, а другие погибают от засухи? Н. Фитол. 178 , 719–739 (2008).
Артикул Google ученый
Seybold, S.J. et al. Борьба с короедом западной части Северной Америки с помощью семиохимических препаратов. Год. Преподобный Энтомол. 63 , 407–432 (2018).
КАС пабмед Статья Google ученый
Fettig, CJ, McKelvey, S.R. & Huber, D.P.W. Летучие вещества покрытосеменных, не являющиеся хозяевами, и вербенон нарушают реакцию западного соснового лубоеда, Dendroctonus brevicomis (Coleoptera: Scolytidae), на ловушки с приманкой-аттрактантом.
Ж. эконом. Энтомол. 98 , 2041–2048 (2005).КАС пабмед Статья Google ученый
Fettig, CJ, Dabney, C.P., McKelvey, S.R. & Huber, D.P.W. Летучие вещества покрытосеменных, не являющиеся хозяевами, и вербенон защищают отдельные сосны пондерозы от нападения западного соснового жука и красного скипидарного жука (Coleoptera: Curculionidae, Scolytinae). West J. Appl. 23 , 40–45 (2008).
Google ученый
Fettig, C.J. et al. Эффективность препарата «Вербенон Плюс» для защиты сосны желтой и древостоев с 9 лет.0604 Нападение Dendroctonus brevicomis (Coleoptera: Curculionidae) в Британской Колумбии и Калифорнии. Ж. эконом. Энтомол. 105 , 1668–1680 (2012).
ПабМед Статья Google ученый
Fettig, C. & McKelvey, S. Устойчивость внутренних сосновых лесов пондероза к заражению короедом после снижения расхода топлива и лесовосстановительных обработок. Леса 5 , 153–176 (2014).
Артикул Google ученый
Феттиг, К. Дж. и Хильщаньски, Дж. Короеды 555–584. https://doi.org/10.1016/B978-0-12-417156-5.00014-9 (Эльзевир, 2015 г.).
Чессон, П. Механизмы поддержания видового разнообразия. Год. Преподобный Экол. Сист. 31 , 343–366 (2000).
Артикул Google ученый
Fricker, G. A. et al. Больше, чем климат? Предикторы высоты кроны деревьев варьируются в зависимости от масштаба сложной местности, Сьерра-Невада, Калифорния (США).
Для. Экол. Управление 434 , 142–153 (2019).Артикул Google ученый
Ма, С., Консилио, А., Окли, Б., Норт, М. и Чен, Дж. Пространственная изменчивость микроклимата в смешанном хвойном лесу до и после рубок ухода и выжигания. Для. Экол. Управление 259 , 904–915 (2010).
Артикул Google ученый
Stovall, A.E.L., Shugart, H. & Yang, X. Высота дерева объясняет риск гибели во время сильной засухи. Нац. коммун. 10 , 1–6 (2019).
Артикул КАС Google ученый
Stephenson, N. L. & Das, A. J. Изменения в составе леса, связанные с высотой, объясняют увеличение смертности деревьев с высотой во время сильной засухи. Нац. коммун. 11 , 3402 (2020).
ОБЪЯВЛЕНИЕ КАС пабмед ПабМед Центральный Статья Google ученый
Человек, Х. Л. Селекция деревьев западным сосновым жуком. Дж. Для. 26 , 564–578 (1928).
Google ученый
Person, HL Теория, объясняющая выбор определенных деревьев западным сосновым жуком. Дж. Для. 29 , 696–699 (1931).
КАС Google ученый
Пайл, Л. С., Мейер, М. Д., Рохас, Р., Роу, О. и Смит, М. Т. Воздействие засухи и совокупная смертность лесных деревьев в южной части Сьерра-Невады. Леса 10 , 237 (2019).
Артикул Google ученый
Фрей Дж., Ковач К., Штеммлер С. и Кох Б. Фотограмметрия лесов с БПЛА как уязвимый процесс. Анализ чувствительности структуры из конвейера RGB-изображения движения. Дистанционный датчик 10 , 912 (2018 г.).
ОБЪЯВЛЕНИЕ Статья Google ученый
Джеймс, М. Р. и Робсон, С. Уменьшение систематической ошибки в топографических моделях, полученных с помощью БПЛА и наземных сетей изображений. Прибой Земли. Процесс. Ландф. 39 , 1413–1420 (2014).
ОБЪЯВЛЕНИЕ Статья Google ученый
Грей, П. К. и др. Сверточная нейронная сеть для обнаружения морских черепах на снимках с дронов. Методы Экол. Эвол. 10 , 345–355 (2019).
Артикул Google ученый
Восе, Дж. М. и др. в I воздействия, риски и адаптация в Соединенных Штатах: Четвертая национальная оценка климата , том II (ред. Рейдмиллер, Д. Р. и др.) 232–267. https://nca2018.globalchange.gov/chapter/6/https://doi.org/10.7930/NCA4.2018.CH6 (2018).
Bedard, W.D. et al. Западный сосновый жук: полевая реакция на его половой феромон и синергический терпен-хозяин, мирцен. Наука 164 , 1284–1285 (1969).
ОБЪЯВЛЕНИЕ КАС пабмед Статья Google ученый
Байерс, Дж. А. и Вуд, Д. Л. Межвидовое ингибирование реакции жуков-короедов, Dendroctonus brevicomis и Ips paraconfusus , к их феромонам в полевых условиях.
J. Chem. Экол. 6 , 149–164 (1980).КАС Статья Google ученый
Шеперд, В. П., Хубер, Д. П. В., Сейболд, С. Дж. и Феттиг, С. Дж. Реакция усиков западного соснового лубоеда, Dendroctonus brevicomis (Coleoptera: Curculionidae), на стеблевые летучие вещества его основного хозяина, 6 09 Pinus ponderosa и девять симпатрических нехозяин покрытосеменных и хвойных. Химиоэкология 17 , 209–221 (2007).
КАС Статья Google ученый
DJI. Zenmuse X3 — свобода творчества. Официальный DJI https://www.dji.com/zenmuse-x3/info (2015).
Микасенс. МикаСенс. https://support.micasense.com/hc/en-us/articles/215261448-RedEdge-User-Manual-PDF-Download- (2015 г.).
DJI. DJI — мировой лидер в производстве дронов/квадрокоптеров для аэрофотосъемки.
Официальный номер DJI https://www.dji.com/matrice100/info (2015 г.).Wyngaard, J. et al. Возникающие проблемы для управления научными данными sUAS: справедливость за счет участия сообщества и разработки передового опыта. Дистанционный датчик 11 , 1797 (2019).
ОБЪЯВЛЕНИЕ Статья Google ученый
Роуз В., Хаас Р. Х., Диринг В. и Шелл Дж. А. Мониторинг весеннего продвижения и ретроградации (эффект зеленой волны) естественной растительности (Центр дистанционного зондирования, Техасский университет A&M, 1973 г.).
DronesMadeEasy. Map Pilot для DJI на iOS. Магазин приложений https://itunes.apple.com/us/app/map-pilot-for-dji/id1014765000?mt=8 (2018 г.).
Farr, T.G. et al. Топографическая миссия шаттла. Ред. Геофиз. 45 , RG2004 (2007 г.
).Zhang, W. et al. Простой в использовании метод фильтрации бортовых данных LiDAR, основанный на моделировании ткани. Дистанционный датчик 8 , 501 (2016).
ОБЪЯВЛЕНИЕ Статья Google ученый
Hijmans, R.J. et al. Растр: анализ географических данных и моделирование (2019).
Гительсон А., Мерзляк М. Н. Изменения спектральной отражательной способности, связанные с осенним старением листьев Aesculus hippocastanum L. и Acer platanoides L.. Спектральные особенности и связь с оценкой хлорофилла. J. Физиол растений. 143 , 286–292 (1994).
КАС Статья Google ученый
Купс, Н. К., Джонсон, М., Вулдер, М. А. и Уайт, Дж. К. Оценка изображений с высоким пространственным разрешением QuickBird для обнаружения повреждений, вызванных атакой красного цвета из-за заражения горным сосновым жуком.
Дистанционный датчик окружающей среды. 103 , 67–80 (2006).ОБЪЯВЛЕНИЕ Статья Google ученый
Клеверс, Дж. Г. П. В. и Гительсон, А. А. Дистанционная оценка содержания хлорофилла и азота в культурах и травах с использованием полос красного края на Sentinel-2 и -3. Междунар. Дж. Заявл. Наблюдение за Землей. Геоинф. 23 , 344–351 (2013).
ОБЪЯВЛЕНИЕ Статья Google ученый
Ли В., Го К., Якубовски М. К. и Келли М. Новый метод сегментации отдельных деревьев из облака точек LiDAR. Фотограмм. англ. Remote Sens. 78 , 75–84 (2012).
Артикул Google ученый
Якубовски М.К., Ли В., Го К. и Келли М. Выделение отдельных деревьев по данным LiDAR: сравнение векторного и растрового подходов к сегментации.
Дистанционный датчик 5 , 4163–4186 (2013 г.).ОБЪЯВЛЕНИЕ Статья Google ученый
Шин, П., Санки, Т., Мур, М. и Тоде, А. Оценка изображений беспилотных летательных аппаратов для оценки топлива в пологе леса в древостое сосны желтой. Дистанционный датчик 10 , 1266 (2018 г.).
ОБЪЯВЛЕНИЕ Статья Google ученый
Руссель Ж.-Р. lidRplugins: дополнительные функции и алгоритмы для пакета lidR (2019).
Эйсн, Л. и др. Эталонный тест методов обнаружения одиночных деревьев на основе LiDAR с использованием разнородных данных о лесах из альпийского пространства. Леса 6 , 1721–1747 (2015).
Артикул Google ученый
Вега, К. и др. PTree: точечный подход к извлечению лесных деревьев из данных LiDAR.
Междунар. Дж. Заявл. Наблюдение за Землей. Геоинф. 33 , 98–108 (2014).ОБЪЯВЛЕНИЕ Статья Google ученый
Плаурайт, А. ForestTools: анализ данных дистанционного зондирования леса (2018).
Пау Г., Фукс Ф., Скляр О., Бутрос М. и Хубер В. EBImage: R-пакет для обработки изображений с приложениями к клеточным фенотипам. Биоинформатика 26 , 979–981 (2010).
КАС пабмед ПабМед Центральный Статья Google ученый
Мейер, Ф. и Бойхер, С. Морфологическая сегментация. Дж. Виз. коммун. Изображение Представлять. 1 , 21–46 (1990).
Артикул Google ученый
Ханзикер, стр. Velox: быстрое извлечение и обработка растра (2017).
Кун М. Построение прогностических моделей в R с использованием пакета Caret. J. Стат. ПО 28 , 1–26 (2008).
Артикул Google ученый
Wang, Y. et al. Является ли высота деревьев, измеренная в полевых условиях, такой же надежной, как считается – сравнительное исследование оценок высоты деревьев, полученных на основе полевых измерений, бортового лазерного сканирования и наземного лазерного сканирования в бореальном лесу. ISPRS J. Фотограмм. Remote Sens. 147 , 132–145 (2019).
ОБЪЯВЛЕНИЕ Статья Google ученый
Вайнштейн, Б.Г., Маркони, С., Болман, С., Заре, А. и Уайт, Э. Обнаружение отдельных крон деревьев на изображениях RGB с использованием полууправляемых нейронных сетей глубокого обучения. Дистанционный датчик 11 , 1309 (2019).
ОБЪЯВЛЕНИЕ Статья Google ученый
душ Сантос, А. А. и др. Оценка методов обнаружения отдельных деревьев на основе CNN на изображениях, снятых RGB-камерами, прикрепленными к БПЛА. Датчики (Базель) 19 , 3595 (2019).
Стивенсон, Н. Фактическая эвапотранспирация и дефицит: биологически значимые корреляты распределения растительности в пространственных масштабах. Ж. Биогеогр. 25 , 855–870 (1998).
Артикул Google ученый
Флинт, Л. Э., Флинт, А. Л., Торн, Дж. Х. и Бойнтон, Р. Мелкомасштабное гидрологическое моделирование для приложений регионального ландшафта: разработка и работа модели характеристики бассейна Калифорнии. Экол. Процесс. 2 , 25 (2013).
Артикул Google ученый
Baldwin, B.G. et al. Видовое богатство и эндемизм местной флоры Калифорнии. утра. Дж. Бот. 104 , 487–501 (2017).
ПабМед Статья Google ученый
Бюркнер, П.-К. brms: пакет R для байесовских многоуровневых моделей с использованием Stan. J. Стат. ПО 80 , 1–28 (2017).
Артикул Google ученый
Хоффман, М. Д. и Гельман, А. Пробоотборник без разворота: адаптивная установка длины пути в гамильтоновом методе Монте-Карло.
Дж. Маха. Учиться. Рез. 15 , 31 (2014).MathSciNet МАТЕМАТИКА Google ученый
Карпентер Б. и др. Стэн: вероятностный язык программирования. J. Стат. ПО 76 , 1–32 (2017).
Артикул Google ученый
Брукс С.П. и Гельман А. Общие методы контроля сходимости итеративного моделирования. Дж. Вычисл. График Стат. 7 , 434 (1998).
MathSciNet Google ученый
Габри Дж., Симпсон Д., Вехтари А., Бетанкур М. и Гельман А. Визуализация в байесовском рабочем процессе. JR Stat. соц. сер. А 182 , 389–402 (2019).
MathSciNet Статья Google ученый
Кунц, М. Дж., Латимер, А. М., Мортенсон, Л.
А., Феттиг, С. Дж. и Норт, М. П. Данные, полученные с помощью дронов, подтверждающие «Межмасштабное взаимодействие размера дерева-хозяина и климатического дефицита воды влияет на гибель деревьев, вызванную короедом». https://doi.org/10.17605/OSF.IO/3CWF9 (2020 г.).Болдуин, Б.Г. и др. Основной пространственный файл для местных сосудистых растений Калифорнии, используемый Baldwin et al. (2017 г., амер. J. Bot.), Дриада, набор данных, 2017 г. https://doi.org/10.6078/D16K5W.
R Основная команда. R: язык и среда для статистических вычислений (R Foundation for Statistical Computing, 2018).
Кунц, М.Дж., Латимер, А.М., Мортенсон, Л.А., Феттиг, С.Дж. и Норт, М.П. Локальная структура-wpb-серьезность. https://doi.org/10.17605/OSF.IO/WPK5Z (2019 г.).
 «>
«>Робсон, С. М. Пересмотр недавней засухи в Калифорнии как экстремального значения. Геофиз. Рез. лат. 42 , 6771–6779 (2015).
ОБЪЯВЛЕНИЕ Статья Google ученый
 ).
). pdf (USDAFS, 2017).
pdf (USDAFS, 2017). «>
«>Феттиг, С. Дж., Мортенсон, Л. А., Булаон, Б. М. и Фоулк, П. Б. Гибель деревьев после засухи в центральной и южной части Сьерра-Невады, Калифорния, США For. Экол. Управление 432 , 164–178 (2019).
Артикул Google ученый
 J. Appl Ecol. 53 , 530–540 (2016).
J. Appl Ecol. 53 , 530–540 (2016). «>
«>Валлин, К. Ф. и Раффа, К. Ф. Обратная связь между индивидуальным поведением при выборе хозяина и динамикой популяции извергающегося травоядного. Экол. моногр. 74 , 101–116 (2004).
Артикул Google ученый
 «>
«>ДеРоуз, Р. Дж. и Лонг, Дж. Н. История нарушений, вызванных засухой, характеризует субальпийский лес на юге Скалистых гор. Кан. Дж. Рез. 42 , 1649–1660 (2012).
Артикул Google ученый
 Экогидрология 6 , 444–454 (2013).
Экогидрология 6 , 444–454 (2013). «>
«>Thistle, H.W. et al. Суррогатные шлейфы феромонов в трех участках ствола леса: составная статистика и тематические исследования. Для. науч. 50 , 610–625 (2004).
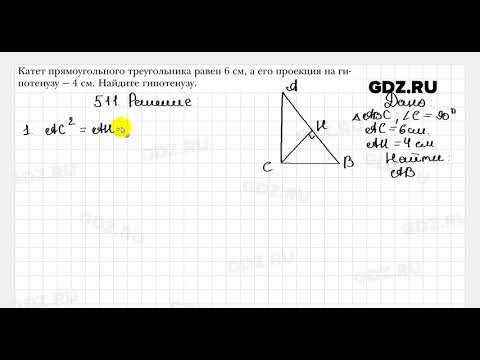 «>
«>Гейзлер, Д. Р. и Гара, Р. И. в «Теория и практика управления горным сосновым жуком в сосновых лесах: материалы симпозиума » (под ред. Берримана, А. А., Аммана, Г. Д. и Старка, Р. В.) (1978).

 Для. Экол. Управление 238 , 24–53 (2007).
Для. Экол. Управление 238 , 24–53 (2007). Кан. Энтомол. 114 , 797–810 (1982).
Кан. Энтомол. 114 , 797–810 (1982). Для. Экол. Управление 267 , 74–92 (2012).
Для. Экол. Управление 267 , 74–92 (2012). «>
«>Руссель, Дж.-Р., Оти, Д., Де Буасье, Ф. и Меадор, А. С. lidR: Бортовая лидарная обработка и визуализация данных для лесохозяйственных приложений (2019).
 Ж. эконом. Энтомол. 98 , 2041–2048 (2005).
Ж. эконом. Энтомол. 98 , 2041–2048 (2005).Oliver, W. W. Управляет ли самопрореживание сосны пондероза короедом Dendroctonus ? В Оздоровление лесов через лесоводство: материалы Национального семинара по лесоводству 1995 г. 6 (1995).
Stovall, A.E.L., Shugart, H.H. & Yang, X. Ответ на «Изменения в составе леса, связанные с высотой, объясняют увеличение смертности деревьев с высотой во время сильной засухи». Нац. коммун. 11 , 3401 (2020).
ОБЪЯВЛЕНИЕ КАС пабмед ПабМед Центральный Статья Google ученый
Миллар С.И., Стефенсон Н.Л. и Стивенс С.Л. Изменение климата и леса будущего: управление в условиях неопределенности. Экол. заявл. 17 , 2145–2151 (2007).
ПабМед Статья Google ученый
Millar, C.I. et al. Смертность лесов в высокогорных белокорых сосновых ( Pinus albicaulis ) лесах восточной Калифорнии, США: влияние экологического контекста, короеды, климатический дефицит воды и потепление. Кан. Дж. Для. Рез. 42 , 749–765 (2012).
Артикул Google ученый
Загрузить ссылки
Благодарности
Мы с благодарностью признаем финансирование от Центра оценки экологических угроз Западных диких земель Министерства сельского хозяйства США (WWETAC) и Программы конкурсных грантов Тихоокеанской юго-западной исследовательской станции по изменению климата. Мы благодарим Конни Миллар за комментарии и рекомендации во время разработки этого проекта, а также Миган Олдфатер за ее роль визуального наблюдателя во время полетов дронов. Мы благодарим Беверли Булаон за ее значительный вклад в создание сети полевых участков. Мы также благодарим Викторию Шолль за полезные обсуждения уровней продуктов данных дистанционного зондирования и Дерека Янга за полезные обсуждения при редактировании этой рукописи. Мы благодарим команду Map Pilot для iOS (которая реализовала несколько запросов на функции, которые помогли нам провести наши полеты дронов), Pix4D (предоставившую бесплатную облачную инфраструктуру для большей части обработки фотограмметрии Structure from Motion) и Open Science Framework (которая помогла публичный доступ к нашему полному набору данных). Спасибо Алексу Манделю, Дэну Крофчеку, Тейлору Нельсону, Нейту Мецлеру, Брэндону Старку, Энди Вонгу, Грейс Лю, Шону Хогану, команде Micasense, Лоуренсу Деннису из Aerial Technology International и Кейси Нейстат за ценный вклад в отношении дронов, датчиков и безопасных полетов. и фотограмметрии SfM. Публикация этой статьи частично финансировалась Фондом открытого доступа библиотек Университета Колорадо в Боулдере и Фондом открытого доступа библиотеки Дэвиса Калифорнийского университета.
Информация об авторе
Авторы и организации
Выпускная группа по экологии, Калифорнийский университет, Дэвис, Калифорния, США
Майкл Дж. Кунц, Эндрю М. Латимер и Малкольм П. Норт
3
- 6 90 Департамент завода наук, Калифорнийский университет, Дэвис, Калифорния, США
Майкл Дж. Кунц, Эндрю М. Латимер и Малкольм П. Норт
Earth Lab, Колорадский университет в Боулдере, Боулдер, Колорадо, США
Майкл Дж. Кунц
Лесная служба Министерства сельского хозяйства США, Тихоокеанская юго-западная исследовательская станция, Плейсервилль, Калифорния, США
Лейф А. Мортенсон
Лесная служба Министерства сельского хозяйства США, Тихоокеанская юго-западная исследовательская станция, Дэвис, Калифорния, США
3 Кристофер 90 J.
FettigЛесная служба Министерства сельского хозяйства США, Pacific Southwest Research Station, Mammoth Lakes, CA, США
Malcolm P. North
Авторы
- Michael J. Koontz
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Эндрю М. Латимер
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Leif A. Mortenson
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Кристофер Дж. Феттиг
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Malcolm P. North
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Вклады
Вклад авторов определяется с помощью таксономии ролей участников (CRediT; https://casrai. org/credit/). Концептуализация: MJK, AML, CJF, MPN и LAM; курирование данных: MJK; формальный анализ: MJK; получение финансирования: MJK, MPN, CJF и AML; расследование: MJK, LAM и CJF; методология: M.J.K. и АМЛ; администрирование проекта: M.J.K., MPN и A.M.L.; ресурсы: MJK, MPN и AML; программное обеспечение: M.J.K.; надзор: MJK, MPN и AML; проверка: M.J.K.; визуализация: M.J.K.; написание — первоначальный вариант: MJK; написание — рецензирование и редактирование: M.J.K., A.M.L., C.J.F., MPN и L.A.M.
Автор, ответственный за переписку
Переписка с Майкл Дж. Кунц.
Заявление об этике
Конкурирующие интересы
Авторы не заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Информация о рецензировании Nature Communications благодарит Майкла Райана и других анонимных рецензентов за их вклад в рецензирование этой работы. Доступны отчеты рецензентов.
Примечание издателя Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Дополнительная информация
Дополнительная информация
Файл рецензирования
Права и разрешения
Открытый доступ Использование, распространение, распространение этой статьи под лицензией Creative Commons. и воспроизведение на любом носителе или в любом формате, при условии, что вы укажете автора(ов) оригинала и источник, предоставите ссылку на лицензию Creative Commons и укажете, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons для статьи, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или выходит за рамки разрешенного использования, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатка и разрешения
Об этой статье
Дополнительная литература
Мониторинг лесов в масштабах всей Европы подтверждает необходимость совместной природоохранной стратегии для спасения видов европейского ясеня (Fraxinus spp.
)- Ян-Питер Джордж
- Таня Г. М. Сандерс
- Мейт Ланг
Научные отчеты (2022)
Политика и экология лесного смягчения последствий изменения климата: вызовы, потребности и возможности
- Кортни Л. Гибинк
- Грант М. Домке
- Маргарет Э. К. Эванс
Растения и почва (2022)
- Кортни Л.
Влияние температуры на передвижение и пищевое поведение крупного жука-люпина Sitona gressorius
- Сара Ханниган
- Клаас Нендель
- Маркос Крулл
Journal of Pest Science (2022)
Комментарии
Отправляя комментарий, вы соглашаетесь соблюдать наши Условия и Правила сообщества.